

2025-06-23
研究物种:艾
发表期刊:Plant Biotechnology
发表时间:2022年6月
影响因子:9.803
研究背景
艾与青蒿同样著名,是蒿属中具有巨大经济价值的药用植物,蒿属是菊科中的一个大的植物属,包括大约500个物种和亚种,主要分布在北半球温带地区,被广泛用于药草、食品、化妆品、香料、草料和观赏植物等各个领域。2015年,从青蒿中分离出的抗疟成分青蒿素的发现获得诺贝尔生理学或医学奖,引起全球关注青蒿属其他物种的研究。
细胞遗传学研究促进了对蒿属内物种的系统分类和进化关系的认知,但染色体融合在蒿属基本染色体数目变异中的作用尚未得到充分验证。药理学研究表明,艾表现出抗炎作用、抗过敏、抗菌的、抗氧化剂和抗癌等活性,是由于其叶片中含有大量的次级代谢产物,包括挥发油、黄酮类、萜类化合物、酚酸和其他化合物。
然而,由于缺乏艾高质量的参考基因组,艾的进化和大量活性成分的生物合成的分子基础很少被报道,将为探索艾和其他蒿属植物的遗传和进化生物学提供宝贵的资源。
研究方法
基于Illumina、PacBio HiFi测序和Hi-C相结合完成了艾基因组的组装和注释;后续进行了比较基因组学、转录组学、代谢组等的联合分析。
研究结果
结果1、艾基因组组装及注释
艾染色体级别的基因组组装大小约为8.03 Gb,包含12,499个scaffold,N50大小为206.40 Mb,contig的N50大小为6.25 Mb(图1),Hi-C数据挂载到34条染色体上,挂载率为91.4%。BUSCO完整度95.5%,重复序列比例73.59%。艾基因组共包括93888个基因,基因和编码DNA(CDS)的平均序列长度分别为3416和1256 bp,每个基因平均有5个外显子和4个内含子。此外,这些基因共有93.87%和93.98%在Nr和TrEMBL数据库中被功能注释,72.30%的基因被GO分类,31.13%的基因被注释到KEGG通路。
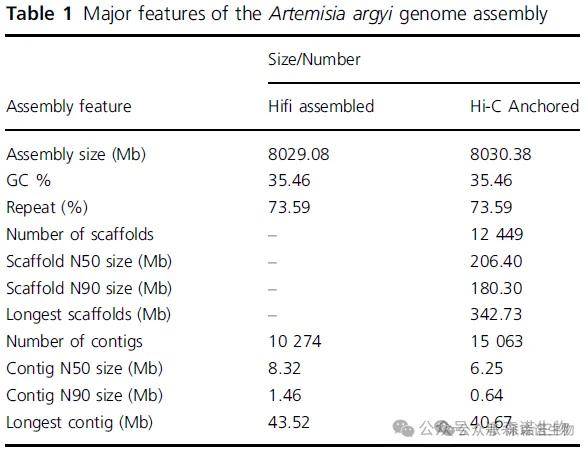
表1. 艾基因组统计
结果2、艾的比较基因组学分析
比较基因组分析鉴定到艾特有的基因家族5321个(包含18,902个特有基因)。在艾基因组中40.04%(11,177/27,915)的基因家族发生扩张,14.15%(3951/27,915)的基因家族收缩(图1B),扩增基因的功能在结合、催化活性、光系统II中的光合电子传递和氧化还原酶活性方面显著相关。这些基因与植物生长和胁迫反应有关,这些大量扩展的基因可能有助于其在整个地球表面的成功扩张和快速生长。通过单拷贝系统发育树和已知化石时间估计出,艾草与青蒿在约740百万年前分化开。WGD分析表明艾经历了两轮WGD(最近的WGD和WGT-1)事件(图2C),最近的WGD事件发生在约2.2百万年前,这是一个物种特异性重复事件,没有发生在短莛飞蓬、向日葵和红花的基因组中,但发生在青蒿和野菊。艾的WGT-1事件是与短莛飞蓬、红花和其他Asterid-II植物共享的保守的全基因组三倍化事件(Badouin 等,2017),发生在大约62.9百万年前。最近的WGD事件和最近在艾中爆发的LTR可能是其基因组大小大的最重要原因之一(图2D)。
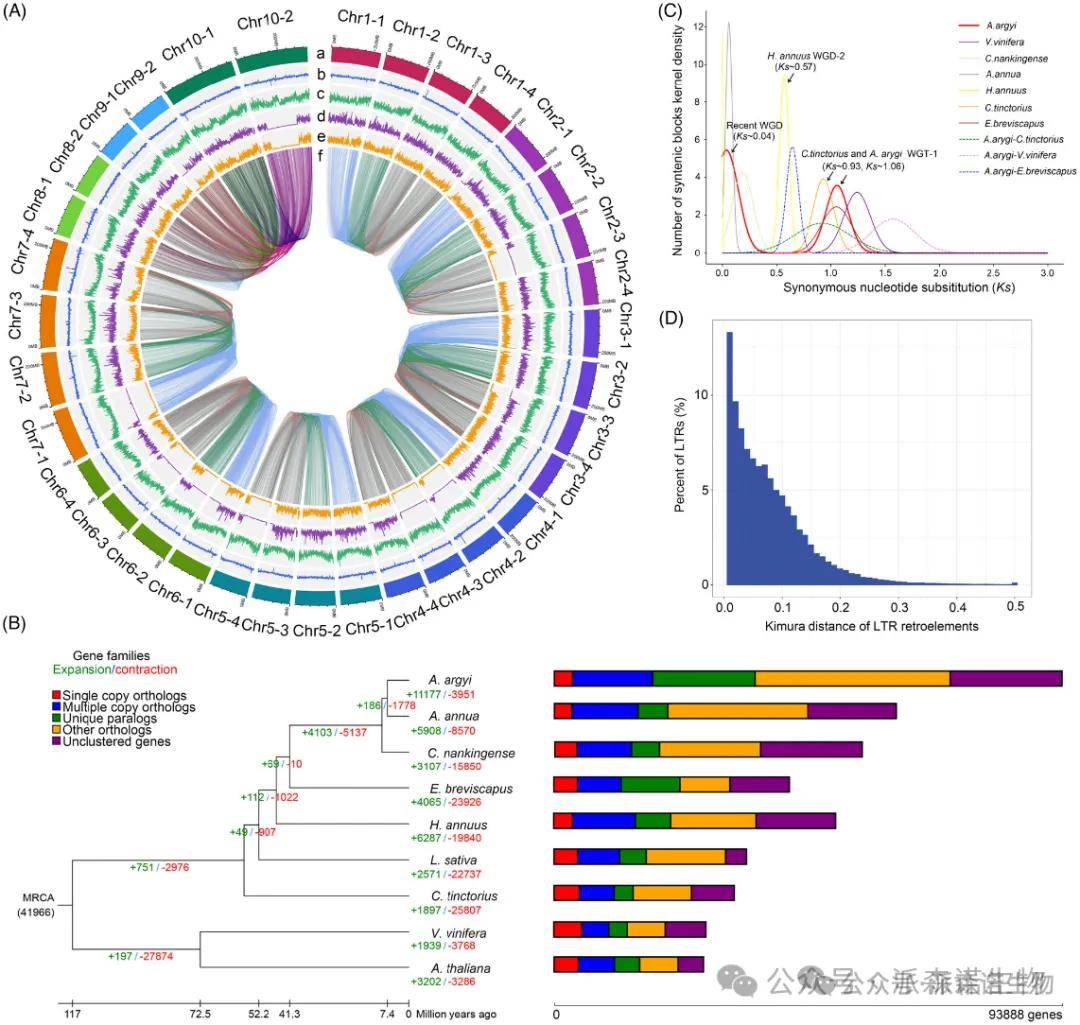
图2. 艾基因组组结果和比较基因组学
结果3、艾基因组的染色体融合
共线性分析表明,艾的34条假染色体由10个同源组组成,其中7个组每组有4套单倍体染色体,3组每组有2条染色体。此外,根据基因共线性分析,1-7染色体组中的四个染色体中的每一个都可以分为两个亚组,这表明艾是异源四倍体,该结果也与基因组调查分析的结果一致。10号染色体的长度几乎是8号和9号染色体长度的总和(图3a),基因组内共线性分析显示,8号和9号染色体与10号染色体共享紧密的共线区域(图3b)。此外,根据BLAST结果,10号染色体上的11,780个(92.05%,总共12,797个)基因与8号和9号染色体上的基因同源。总之,这些数据使我们能够假设祖先的8号和9号染色体被融合到艾的10号染色体中,并且10号染色体表现为祖先8号和9号染色体的末端融合,伴随通过至少一次倒位和两次染色体内易位事件。
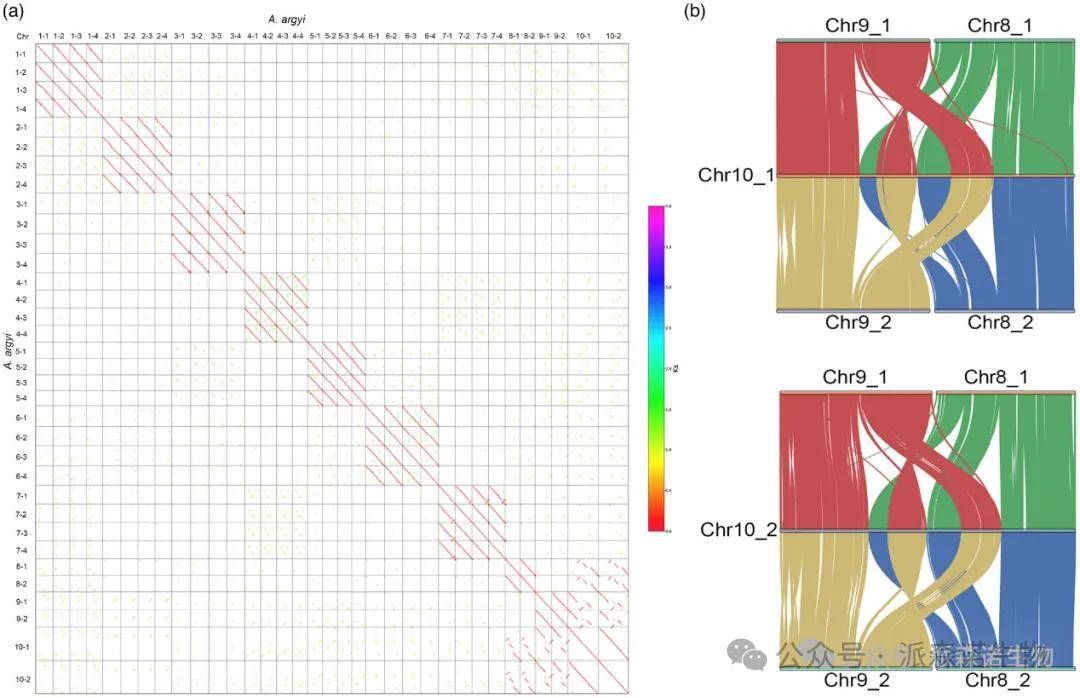
图3. 10号染色体与8号、9号染色体之间的染色体共线性模式和染色质结构。
结果4、参与黄酮合成基因
采用超高效液相色谱(HPLC)对‘香艾’七种不同组织(根、根茎、茎和叶A-D的四个不同发育阶段)中的这些黄酮类化合物进行定量。结果表明,这些生物活性黄酮类化合物在叶子中比在其他组织中更丰富,其中泽兰黄醇素含量最高,且随叶片生长期而增加。基于在其他物种中对黄酮的广泛研究,我们提出了这些化合物在艾中可能的生物合成途径(图4a,c)。
通过同源搜索和功能注释,在黄酮类生物合成途径中共鉴定出44个候选基因共编码12种关键酶,近一半的候选基因属于扩张基因。利用来自根和叶器官的样本进行转录组分析,以鉴定不同组织和不同发育阶段的叶片之间的差异表达基因。几乎所有候选基因都在七个选定的组织中都表达,但在该途径中前四个基因的表达水平,特别是根样品中HCT基因的表达水平高于叶片样品,而下游基因,尤其是CHS基因在叶片中的表达水平高于在根中的表达水平(图4b)。
使用保守结构域和已报道的FOMT基因家族作为参考,对艾中的FOMT基因家族进行鉴定与表达模式分析。在艾基因组中找到83个FOMT基因,根据系统发育分析结果划分为五个主要亚群,包括31个3’FOMT、6个6FOMT、10个4’FOMT、32个咖啡酰CoA O-甲基转移酶(CCoAOMT)和四个未分类的FOMT(图4d)。
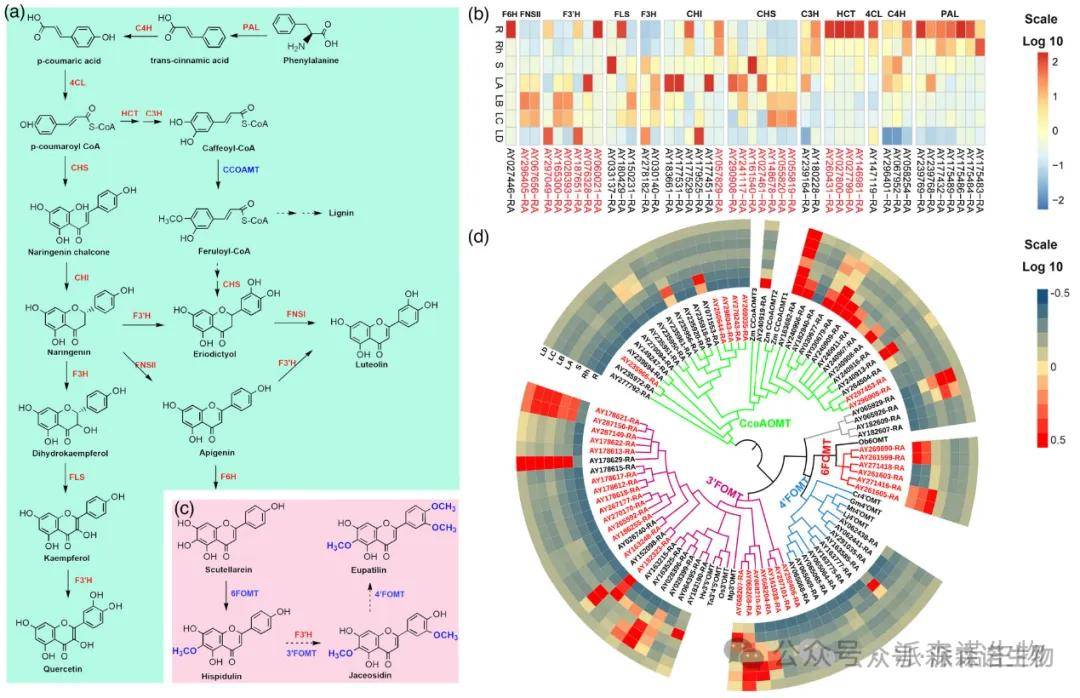
图4. 艾黄酮生物合成相关基因的鉴定和表达谱。
结果5、参与萜类化生物合成基因
由于含有丰富的萜类化合物,挥发油是艾叶中重要的药效学成分。艾叶挥发油中含有丰富的单萜和倍半萜(图5a)。它们都来自常见的前体异戊烯焦磷酸盐(IPP)和二甲基烯丙基焦磷酸盐(DMAPP)。IPP和DMAPP主要通过细胞质中的甲羟戊酸途径(MVA)和质体中的甲基赤藓糖醇磷酸(MEP)途径产生(Sapir-Mir et al., 2008; Vranova et al., 2013)。采用同源搜索和功能注释的方法筛选出参与MVA和MEP通路的候选基因。结果表明,共有66个基因编码14个基因家族,参与了艾的这两个途径(图5b)。
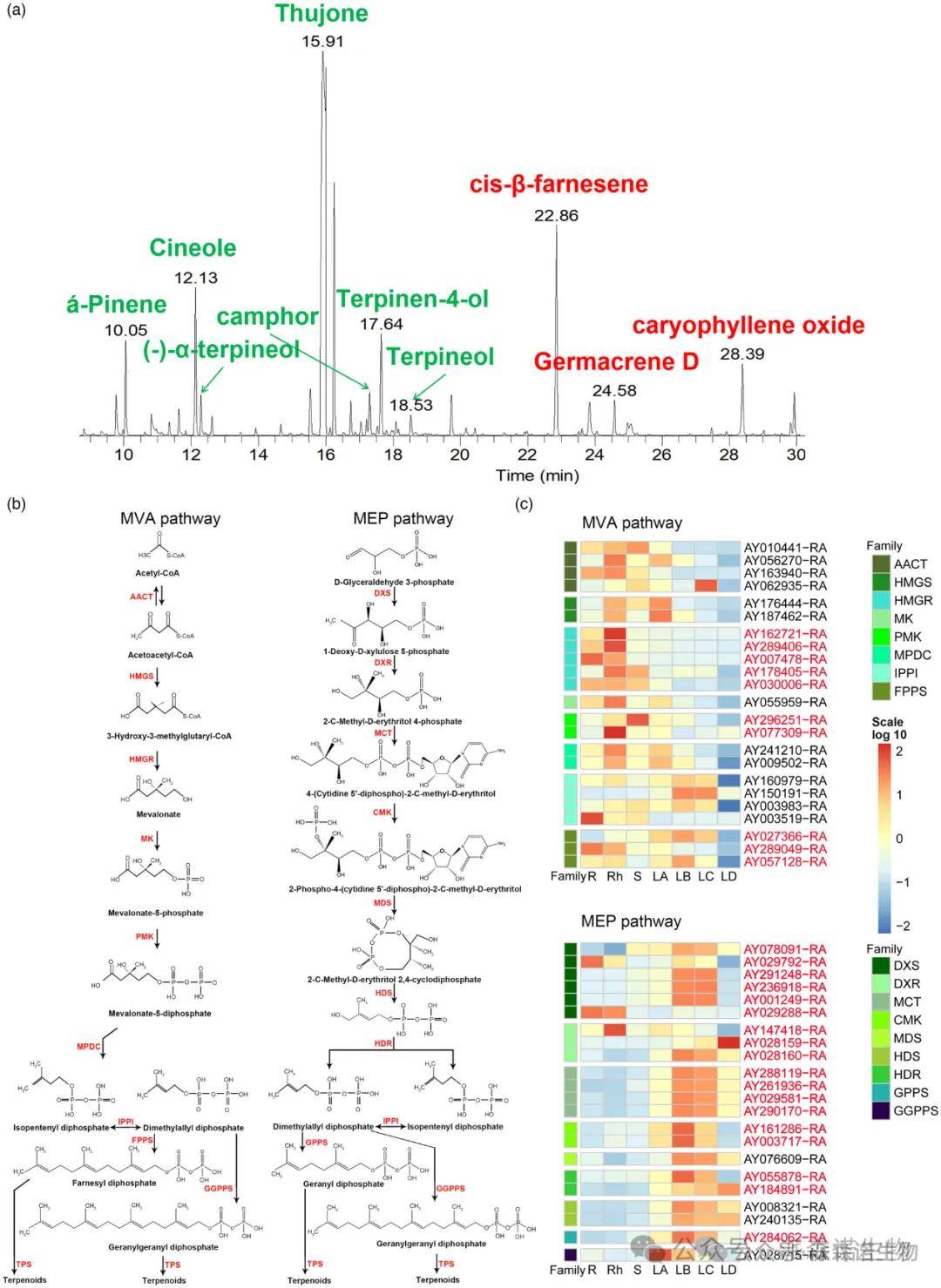
图5. 艾叶中的挥发性化合物及其生物合成途径。
结 论
本项研究报道了蒿属第一个物种艾的染色体级别的参考基因组,基因组带有大量的蛋白编码基因。对艾基因组进行分析,发现艾的3次全基因组复制事件以及近期发生的重复序列的急剧扩张是导致其超大基因组的主要原因。共线性分析结果表明,10号染色体就是蒿属祖先的8、9号两条染色体首尾相连融合后的产物。此外,光系统II中的PsbA基因、DNA复制中的复制因子A1(RPA1)基因、热激蛋白(HSPs)基因和萜类合成酶(TPS)基因极大的扩张,这些基因家族的显著扩张可能是艾具有较强的生态位竞争能力和环境适应性的重要基础。此外,研究还提出了艾黄酮类化合物和萜类化合物的生物合成主干途径,揭示了广泛的基因扩增和串联重复是艾黄酮类化合物和挥发油含量丰富的原因。
参考文献
Yuhuan Miao, Dandan Luo, Tingting Zhao, et al. Genome sequencing reveals chromosome fusion and extensive expansion of genes related to secondary metabolism in Artemisia argyi. Plant Biotechnology. 2022 June 11; https://doi.org/10.1111/pbi.13870


















