

2025-07-03
文章信息
英文题目:Combined GWAS and eGWAS reveals the genetic basis underlying drought tolerance in emmer wheat(Triticum turgidum L.)
中文题目:结合全基因组关联分析和表达数量性状位点分析揭示二粒小麦(Triticum turgidum L.)耐旱性的遗传基础
发表期刊:New Phytologist
影响因子:8.1/Q1
发表时间:2024.01.25
涉及组学:GWAS、转录组测序、eGWAS。
摘要
干旱是全球小麦生产面临的主要环境制约因素之一。作为普通小麦的祖先和基因库,二粒小麦被视为培育耐旱小麦的宝贵基因资源库。
通过对107份种质材料进行GWAS和eGWAS联合分析,我们鉴定出86个数量性状位点(QTL)、105462个表达数量性状位点(eQTL)以及68个与二粒小麦耐旱性(DT)相关的eQTL热点区域。
研究构建了一个由185个上游调控因子和2432个下游干旱响应候选基因组成的复杂调控网络,其中发现TtOTS1基因通过影响根系发育对耐旱性产生负调控作用。
本研究揭示了耐旱性的遗传基础,将为优良耐旱小麦品种的改良育种提供不可或缺的基因资源和种质材料。
前言
小麦(Triticum aestivum L.)是全球最重要的主粮作物之一,其年产量超过6亿吨,为人类提供了约20%的热量摄入。干旱胁迫被认为是限制小麦生产最具破坏性的环境因素。据报道,1980年代至2010年代期间,干旱导致的小麦减产幅度高达21%,严重威胁全球粮食安全。为应对人口增长与气候恶化的双重挑战,急需培育高产耐旱的小麦品种。尽管已有大量研究揭示了调控小麦干旱响应与耐受性的数量性状位点(QTLs)、候选基因及分子模块,但优异基因资源匮乏和遗传多样性狭窄仍是小麦改良面临的突出问题。四倍体二粒小麦(Triticum turgidum L.)作为现代面包小麦A、B亚基因组的祖先种,贡献了小麦三分之二的遗传物质,其基因库包含野生型、地方品种和现代栽培品种。二粒小麦丰富的遗传多样性为提升小麦产量和环境适应性提供了不可或缺的遗传资源。因此,挖掘二粒小麦中控制耐旱性的关键物种特异性基因,有望通过直接杂交或基因工程手段培育超级耐旱小麦。
GWAS与表达数量性状位点(eQTL)分析的结合为解析复杂农艺性状的遗传基础提供了高效手段,已在甘蓝型油菜籽粒含油量、棉花次生细胞壁发育和玉米耐旱性等领域得到应用,但小麦特别是二粒小麦的相关研究仍十分匮乏。本研究通过对107份代表性材料的全基因组重测序数据开展GWAS和eGWAS分析,结合生物量与三个根系相关性状的关联研究,挖掘二粒小麦耐旱性的因果基因并解析其遗传机制。所获eQTLs/QTLs及新候选基因不仅将丰富小麦遗传资源以加速培育超耐旱品种,也为解析耐旱性遗传基础开辟了新途径。
研究内容
1.二粒小麦群体的耐旱性
为评估干旱胁迫下的表现,我们对107份二粒小麦种质资源进行了苗期持续干旱胁迫处理(图1a)。在充分供水(WW)和水分胁迫(WS)条件下测定了各材料的生物量(BM)和根系性状。所有性状在WS条件下均显著低于WW条件(图1b,c),其中生物量在两种水分条件下的变异系数(CV)达33%-38%,较根系性状呈现更大波动。值得注意的是,野生二粒小麦(WEW,圆锥小麦野生亚种)和栽培二粒小麦(DEW,圆锥小麦栽培亚种)的四个性状CV值均因干旱诱导而上升,但硬粒小麦(DW,圆锥小麦硬粒亚种)的CV值却呈现下降趋势(图1d),表明DW在胁迫条件下表现出较低的多样性。
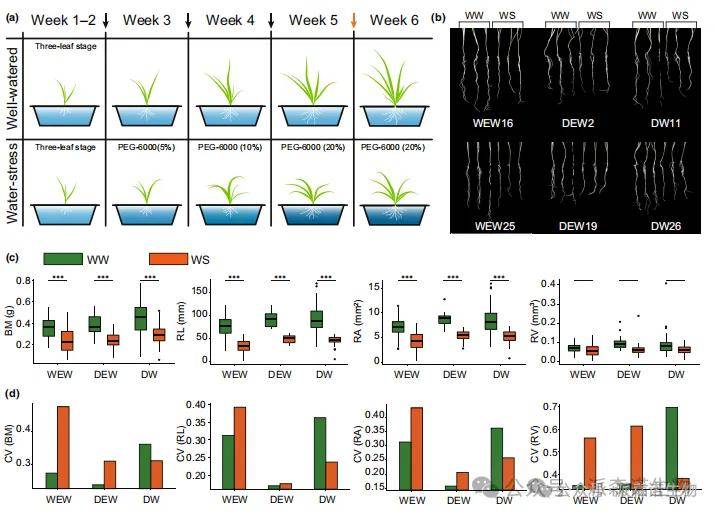
图1.在充分灌溉(WW)和水分胁迫(WS)条件下的二粒小麦表型表现
2.107份二粒小麦种质的群体转录组分析
研究人员进一步对107份种质材料在正常供水(WW)和干旱胁迫(WS)条件下的根组织进行了RNA-Seq分析。主成分分析图将所有样本完全划分为WW和WS两组,表明干旱胁迫引发了显著的转录组差异(图2a)。研究发现根长(RL)与PC1和PC3呈负相关,与PC2呈正相关(图2b),说明根长对表达变异有重要贡献。此外,PC2与生物量(BM)和根表面积(RA)呈正相关,PC3则分别与BM、RA和根体积(RV)呈负相关。
进一步鉴定出9,301个干旱响应基因(图2c),根据其在干旱胁迫下的表达模式可分为三个簇。其中11.1%的基因(簇I)在干旱胁迫下表达下调(图2d),这些基因富集于氧化还原酶活性、蛋白丝氨酸/苏氨酸激酶活性、烟酰胺合成酶活性等蛋白功能条目(图2e)。研究还鉴定出多个已知抗旱基因(图2d),包括Ras相关蛋白(TtRAB18)、小生长素上调基因(TtSAUR1)、脱水素基因(TtXERO1、TtXERO2)、胚胎发育晚期丰富蛋白基因(TtLEA7、TtLEA14、TtLEA29、TtLEA46)以及生长素相关基因(TtJAR1和TtNPH4)。
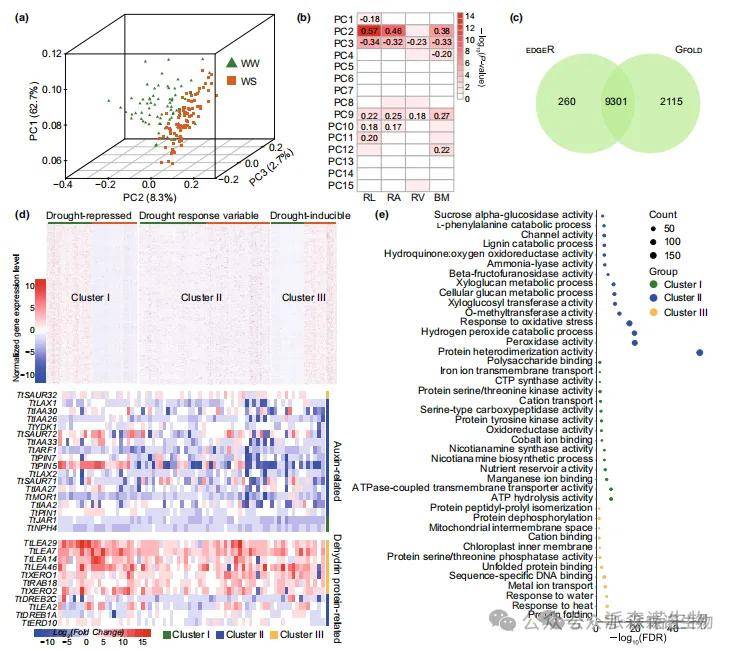
图2.野生二粒小麦响应干旱胁迫的转录组动态变化
3.干旱响应基因表达的选择压力
为研究干旱响应基因表达的自然选择机制,根据干旱胁迫下基因表达丰度与表型性状的关系,计算了单变量线性选择差(S)值。干旱响应基因在BM、RL、RA和RV性状中的平均选择强度(|S|)分别为0.065、0.097、0.070和0.057(图3a),表明大多数基因表达的变异近乎中性。共有4293个独特差异表达基因(DEGs)明显受到四个性状的选择作用(图3b)。其中正向选择基因数量占比超过50%,表现为更高的BM值和更强根系伴随更高基因表达水平。有939个DEGs同时受到四个性状的选择,包括一些既与耐旱性又与根系发育相关的基因(图3c)。例如根系发育相关基因TtMOR1(拟南芥AtMOR1的同源基因)在四个性状中均受到正向选择,平均S值为0.14(图3d)。已被报道与耐旱性相关的AtABA4,在本研究结果中也显示其同源基因TtABA4(拟南芥AtABA4的同源基因)是显著负向选择基因。有趣的是,研究人员还观察到这些选择基因及部分基因家族在三个干旱胁迫下的亚群体表明人工选择也影响了干旱响应基因的表达。
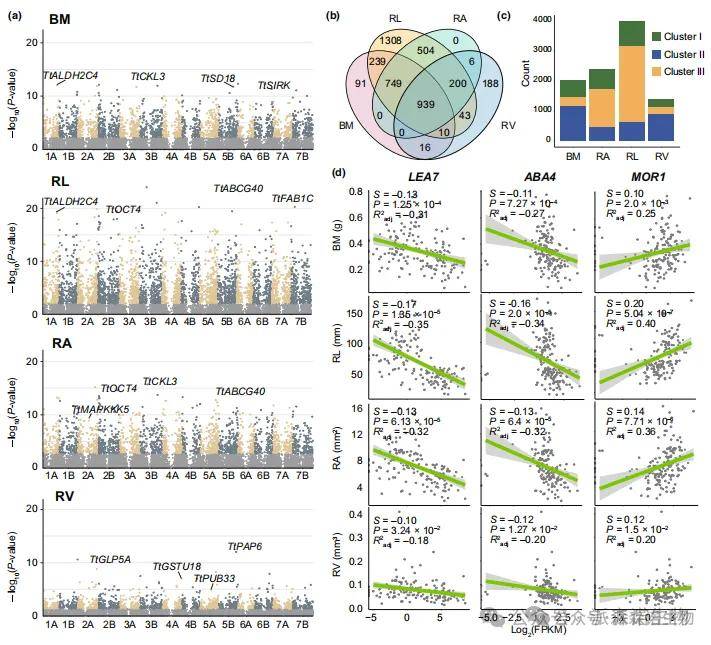
图3.二粒小麦干旱响应基因的选择强度和选择模式
4.二粒小麦耐旱性的eGWAS研究
基于全基因组关联分析(GWAS)采用的相同基因组变异数据集,本研究进一步开展了表达数量性状位点分析(eGWAS),以鉴定与基因表达水平相关的遗传位点。通过分析,共发现105,462个与24,605个eGenes(受eQTL调控的基因)表达相关的eQTLs,其中79.7%的靶向关系通过HiC数据得到验证,证实了所识别eQTLs的准确性。静态eQTLs代表在水分充足(WW)和水分胁迫(WS)条件下均被稳定检测到的eGene,而仅在单一条件下检测到的eQTLs被归类为动态eQTLs。本研究共鉴定出88,664个静态eQTLs和16,798个动态eQTLs(图4a)。结果发现,7.4%的eQTL-eGene对位于同一染色体上(图4b)。此外,研究进一步鉴定出68个eQTL热点区域(包含2366个eQTL位点)(图4c),这些热点调控着8253个基因的表达。每个热点区域调控的eGene数量从177个(Hotspot59)到1674个(Hotspot45)不等。这些受eQTL热点调控的eGene显著富集于干旱胁迫和根系发育相关通路。例如:1B染色体上的Hotspot4调控过氧化物酶及氧化应激响应基因;5A染色体上的Hotspot34调控侧根形成相关基因;5B染色体上的Hotspot42调控过氧化氢分解代谢过程及缺水响应相关基因(图4c)。基于这些eQTL和eGene数据,本研究构建了每个eQTL热点的调控网络。其中Hotspot38网络包含46个静态eQTL和15个动态eQTL,调控1083个下游基因,包括生长素相关基因(TtPIN8、TtIAA8)和转运蛋白基因(TtYSL6、TtABCG17)(图4d);Hotspot51网络包含121个静态eQTL和89个动态eQTL,调控1366个下游基因,其中包括多个转录因子基因(TtbZIP9、TtMYB50、TtWRKY70等)(图4e)。值得注意的是,530个基因位于64个(94.1%)eQTL热点区域内,其中30个编码转录因子或转录调控因子的基因分布在21个eQTL热点中,如TtNAC2、TtMYB14、TtELF6。此外,研究发现4个eQTL热点完全位于基因组的非编码区,提示eQTL中可能还存在调控下游eGene的非编码RNA。
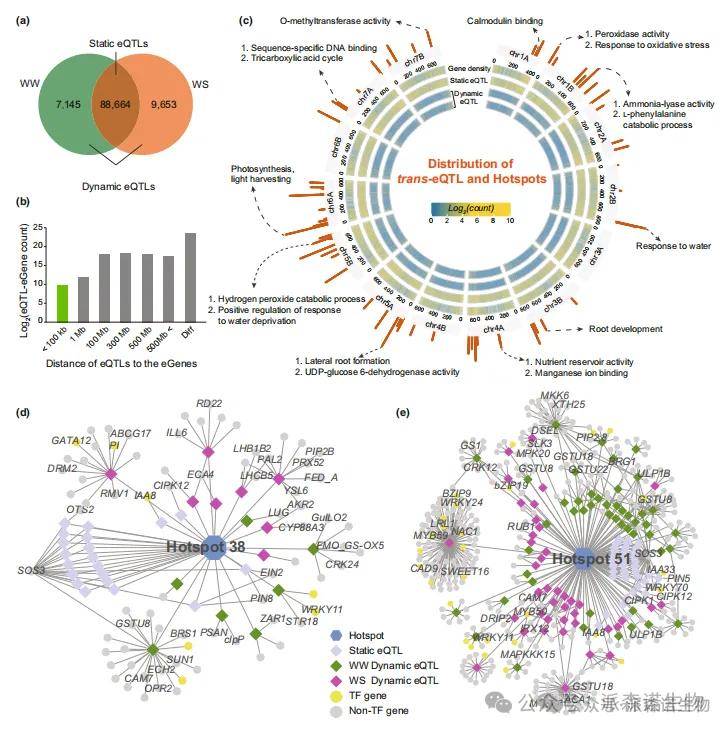
图4.在二粒小麦中鉴定良好灌溉(WW)和水分胁迫(WS)条件下的表达数量性状位点(eQTL)
5.整合数量性状位点与表达数量性状位点揭示抗旱性因果基因
为进一步揭示二粒小麦耐旱性的遗传基础,本研究整合了全基因组关联分析(GWAS)和表达数量性状位点分析(eGWAS)结果。共定位到86个数量性状位点(QTL),包含190个eQTL和989个GWAS信号(图5a,b)。部分变异位点同时与表型性状和基因表达相关。通过二粒小麦与玉米eQTL的共线性分析(Liu et al., 2020),本研究鉴定出18个(9.5%)保守的耐旱相关eQTL,包含TtCDPK、TtUGT85A2、TtSWEETIE等36个上游基因(图5c)。基于共定位的eQTL与QTL,我们进一步构建了二粒小麦耐旱性调控网络。与既往研究一致(Deng et al., 2022),eGenes和区间重叠基因可分为三级:上游层(174个基因)、下游层(5919个基因)和中间层(11个基因),其中包含2432个干旱响应下游基因。上游基因的表达水平显著高于下游基因,且其表达变异性低于中、下游基因。
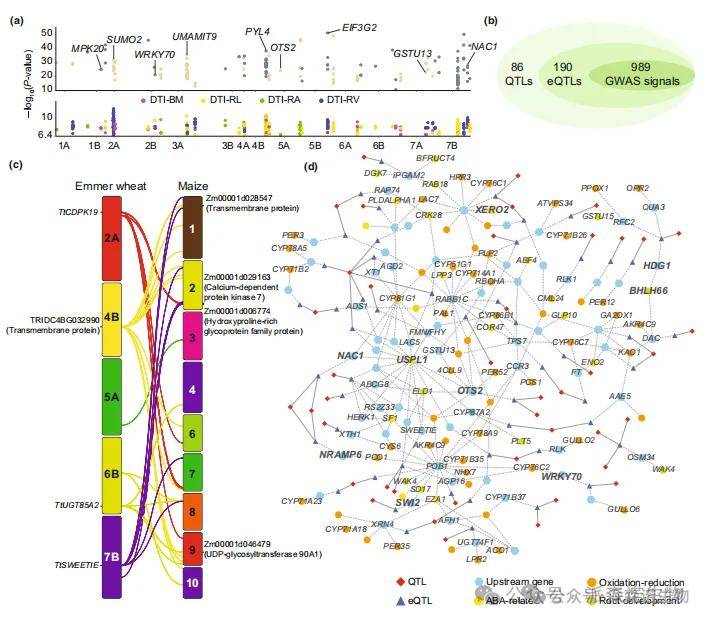
图5.二粒小麦GWAS与eQTL的整合分析
6.TtOTS1是干旱耐受性的负调控因子
基于上述分析,在TRIDC5AG008150.2(反向链)的eQTL chr5A:49280938_49316775(P值=6.31×10^-22)区域内发现一个插入缺失变异(chr5A:49296105,GAGTT>G),该变异与DTIs显著相关(P值=1.96×10^-7)(图6a)。因此,研究人员将其命名为TtOTS1。具有插入基因型的材料(样本数:85)在其CDS序列的第2323 bp处存在AACT序列。TtOTS1基因。相反,缺失基因型(del-AACT)样本(样本数:6)丢失了这四个碱基。该InDel变异位于终止密码子下游13个碱基处,导致移码突变,所有材料可分为ins-AACT单倍型和del-AACT单倍型(图6b)。del-AACT单倍型的DTI值显著大于ins-AACT单倍型(图6c),而其表达水平却低于ins-AACT单倍型(图6d,e),表明其可能作为负调控因子发挥作用。
本研究通过一代测序进一步验证该InDel变异,发现TtOTS1基因的del-AACT变异导致其丢失RRACH基序(R = A/G,H = A/C/U)(图6a),该基序是与m6A甲基化修饰相关的特异性保守基序。为验证该InDel是否影响TtOTS1的m6A甲基化水平,本研究针对变异位点上下游50bp序列进行MeRIP-qPCR检测以排除其他潜在突变位点和RRACH基序干扰。结果显示仅在ins-AACT单倍型材料中检测到m6A修饰(图6f)。此外,TtOTS1作为eQTL(chr4B:418554622_418866532,P值=1.03×10^-50)的eGene,其区间内包含一个m6A阅读蛋白基因(TtECT6),提示m6A修饰可能介导TtOTS1调控二粒小麦的DT性状。
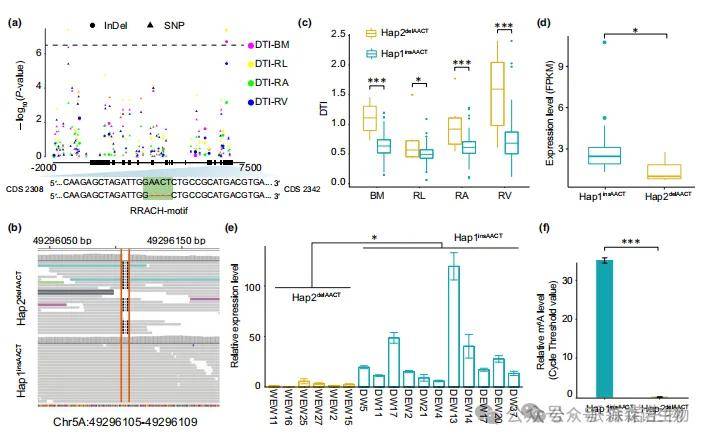
图6 TtOTS1赋予二粒小麦耐旱性
此外,研究团队成功构建了TtOTS1的病毒诱导基因沉默株系(BSMV:OTS)和功能缺失突变株系(ots1),并用于探究其抗旱性。通过定量逆转录聚合酶链式反应和桑格测序技术,验证了VIGS株系和突变株系中TtOTS1表达的下调(图7a,b)。表型分析发现,在干旱条件下,与野生型(WT)相比,TtOTS1沉默植株和翻译提前终止突变体均表现出更高的生物量抗旱指数(DTI-BM)(图7c-e)。进一步研究表明,干旱处理下沉默株和突变株的根长抗旱指数(DTI-RL)显著高于野生型,其根系呈现更发达更深的形态特征(图7f),证实TtOTS1通过调控根系构型负向调控抗旱性。土壤培养实验也显示ots1突变体具有更强的抗旱能力(图S12a,b),再次验证了TtOTS1的功能。
研究进一步对正常供水(WW)和水分胁迫(WS)条件下的VIGS植株进行转录组测序及脱落酸(ABA)含量测定(图7g-i)。共鉴定到3439个差异表达基因(DEGs,其中1700个上调、1739个下调),这些基因显著富集于活性氧(ROS)代谢途径、激素信号转导(ABA和生长素等)及根系发育相关通路。综合研究结果表明,TtOTS1作为二粒小麦抗旱性的负调控因子发挥作用(图7j),该发现为通过基因组编辑技术改良小麦抗旱性提供了潜在靶点。
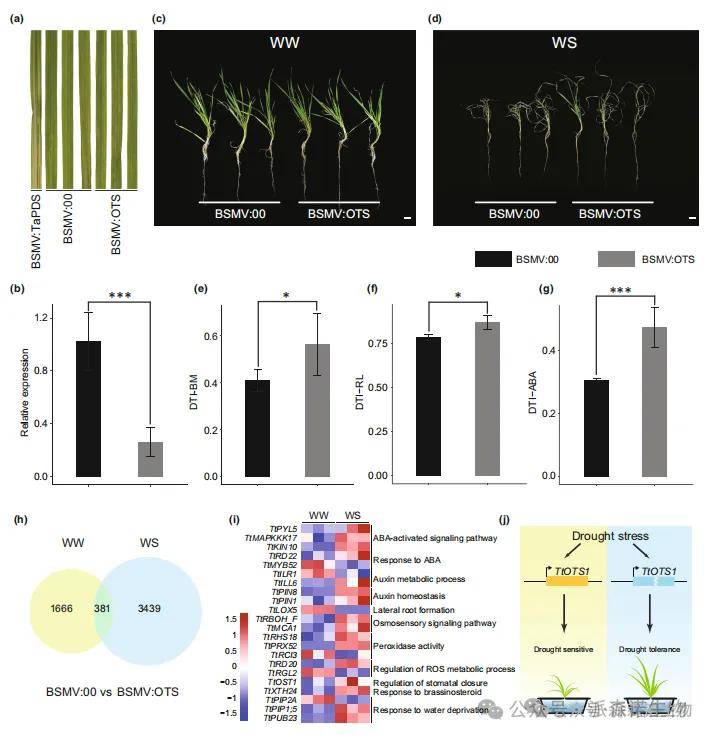
图7 沉默TtOTS1基因增强了二粒小麦的耐旱性。
总结
本研究通过整合全基因组关联分析(GWAS)和表达数量性状位点分析(eGWAS)策略,系统解析了二粒小麦耐旱性(DT)的遗传基础。研究发现107份二粒小麦材料的三个亚群在耐旱指数(DTIs)上无显著差异。然而在水分胁迫(WS)条件下,野生二粒小麦(WEW)材料的变异系数(CV)显著增加,而栽培二粒小麦(DEW)和硬粒小麦(DW)材料变化甚微或有所降低。这些结果表明WEW材料对干旱胁迫的响应具有更高变异性,这与前人研究结果一致。这不仅有助于更好地理解二粒小麦耐旱性的遗传基础,为更好地改良和选育抗旱二粒小麦与面包小麦提供了重要遗传资源。
参考文献:Yang G, Pan Y, Pan W, et al. Combined GWAS and eGWAS reveals the genetic basis underlying drought tolerance in emmer wheat (Triticum turgidum L.). New Phytol. 2024 Jun;242(5):2115-2131. doi: 10.1111/nph.19589.


















