

2025-07-08


文章信息
文章题目:Pangenome analysis reveals structural variation associated with seed size and weight traits in peanut
中文题目:泛基因组分析揭示与花生种子大小和重量相关的结构变异
发表期刊:Nature Genetics
影响因子:29
发表时间:2025年4月
涉及方法:基因组Denovo+全基因组测序+GWAS分析
摘要
花生(Arachis hypogaea L.)是一种重要的油料和食用豆类作物,种子大小和重量是驯化和育种中的关键性状。然而,目前对于种子大小和重量的基因组重排(如结构变异,SVs)的了解仍然有限。本研究通过泛基因组分析,整合了八个高质量基因组(包括两种野生二倍体、两种野生四倍体和四种栽培四倍体花生)以及269份不同种子大小的重测序数据,鉴定出22,222个核心或软核心基因家族、22,232个分布基因家族和5,643个私有基因家族。研究发现,A亚基因组中的SV频率高于B亚基因组,并鉴定出1,335个与驯化相关的SVs和190个与种子大小或重量相关的SVs。这一高质量的泛基因组为花生及其他豆类作物的遗传改良提供了基础资源。
研究背景
花生(Arachis hypogaea L.)是一种重要的油料和食用豆类作物,其种子富含油脂和蛋白质,广泛用于食品工业和食用油生产。种子大小和重量是花生驯化和育种过程中的关键性状,直接影响花生的产量和经济价值。在驯化过程中,花生从野生种的小果品种被选择性地培育为现代的大果品种。然而,目前对于控制这些性状的分子机制和基因组重排(如结构变异,SVs)的了解仍然有限。以往的研究主要集中在SNPs(单核苷酸多态性)和小片段插入/缺失(indels)上,但对于大尺度的基因组重排(如SVs)的研究相对较少。这些SVs可能对种子大小和重量的调控起重要作用。
花生是一种异源四倍体作物,其基因组复杂,包含大量的重复序列和多倍体特性。目前可用的花生基因组资源仍然存在许多缺口,高质量的基因组资源相对匮乏。这主要是由于多倍体基因组的组装和注释难度较大。为了更好地探索基因组变异,需要高质量的基因组资源来覆盖不同种群的花生。泛基因组(pangenome)通过整合来自不同个体或种群的基因组数据,提供了一个全面的视角来更深入地理解物种或种群的基因组结构和功能多样性。
研究结果
1、269份具有不同种子大小材料的遗传多样性分析
对269份具有不同种子大小的花生材料进行了全基因组重测序(总数据量约8 TB),系统评估其遗传多样性和群体结构。这些材料涵盖32份AA二倍体野生种、8份AABB四倍体野生种、155份AABB四倍体地方种、67份AABB四倍体改良品种和7份未分类样本。共获得5,989,854个高质量SNP位点(AA亚基因组:5,048,032个;BB亚基因组:941,822个)。研究发现,AA二倍体野生种的多样性最高(π=8.57×10⁻⁴),而BB二倍体野生种显著较低(π=5.69×10⁻⁵);在四倍体中,BB亚基因组多样性均高于A亚基因组,如AABB地方种在B亚基因组上π=1.46×10⁻⁴,高于A亚基因组的π=1.17×10⁻⁴,显示出B亚基因组在栽培花生中保留更多遗传变异。
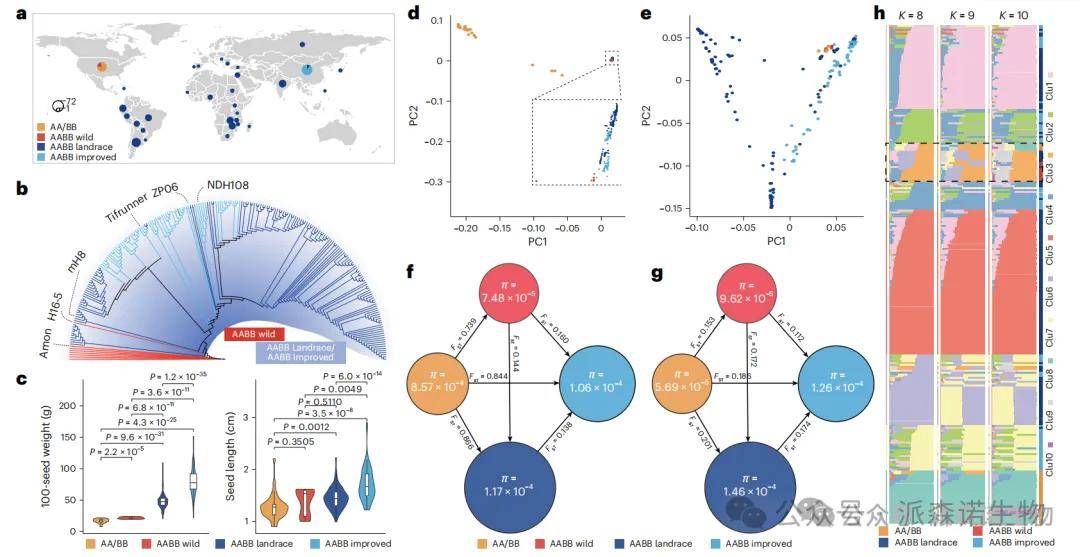
图1 269份野生和栽培花生种质的遗传多样性
2、构建具有多样荚果大小的高质量花生泛基因组
通过整合8个代表性花生材料的高质量基因组,构建了一个结构完整、功能丰富的泛基因组系统。其中,6个新组装基因组使用Nanopore、HiFi和Hi-C技术,组装质量显著提升,scaffold N50达134–148 Mb,BUSCO完整性高达98.6–99.4%。泛基因组共识别50,097个基因家族。新组装基因组平均拥有约37,000个家族,显著多于以往的30,000个(P<0.001),其中1,086个新基因具高表达证据,证明其生物学功能的可靠性。核心和软核心基因在结构上更长、表达量更高、受选择压力更强(P<0.001),广泛参与光合作用、脂质代谢等功能。
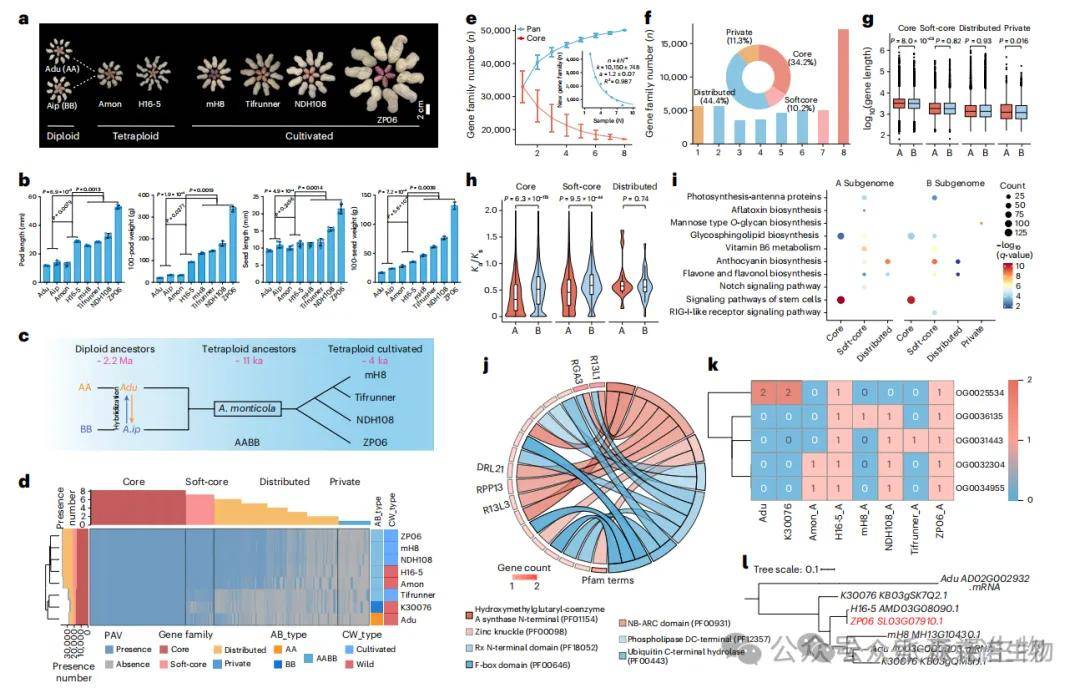
图2 花生的基因水平泛基因组
3、结构变异(SV)可能影响基因表达
通过构建图谱型花生泛基因组,整合组装与比对两种SV检测方法,共识别出86,308个高置信度SV,主要为插入与缺失。其中10–16%的SV直接与基因重叠,15–30%位于启动子或下游3,000 bp区域。启动子和外显子区域中的SV更可能影响基因表达,最多有40%的插入/缺失来源于LTR或DNA转座子。以大荚材料ZP06为例,其A亚基因组包含2,130个插入和2,160个缺失,B亚基因组也发现大量结构变异。多个重要基因如Snakin-2、NAD+合成酶和糖基转移酶等的启动子或外显子中SV改变了其表达量或结构,如7.9 kb缺失、1.2 kb插入等,直接影响产量与抗性。
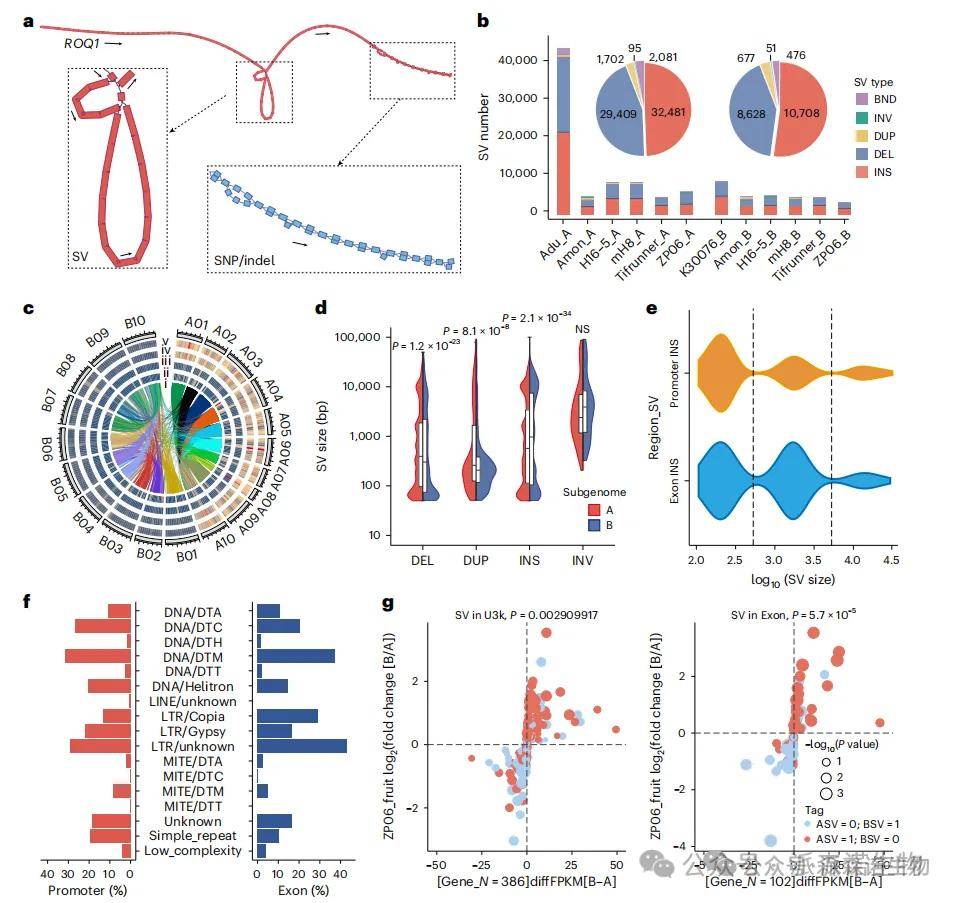
图3 泛基因组中结构变异(SVs)的特征
4、驯化过程中性状相关基因的选择性扫描
研究表明,结构变异(SVs)在花生驯化过程中对果实大小和抗病性状的形成起到了关键作用。在驯化相关SVs中,约41–55%与LTR或Gypsy重复元件重叠,35–55%与简单重复序列重叠,且B亚基因组的Helitron转座子比例显著低于A亚基因组,显示出明显的进化特征。19个SV相关基因与果实大小和抗病性状显著相关。例如,钙依赖性激酶CRK26在多个野生和二倍体品种中存在629 bp缺失,而亮氨酸重复基因IRK中有27.7 kb缺失,通过基因组组装得以准确识别。此外,荚果大小相关基因NTF6与抗病基因FBRL2构成花生中特有的串联单元,其高拷贝数变异与大荚果表型密切相关,且在抗病情境下表达同步下降。
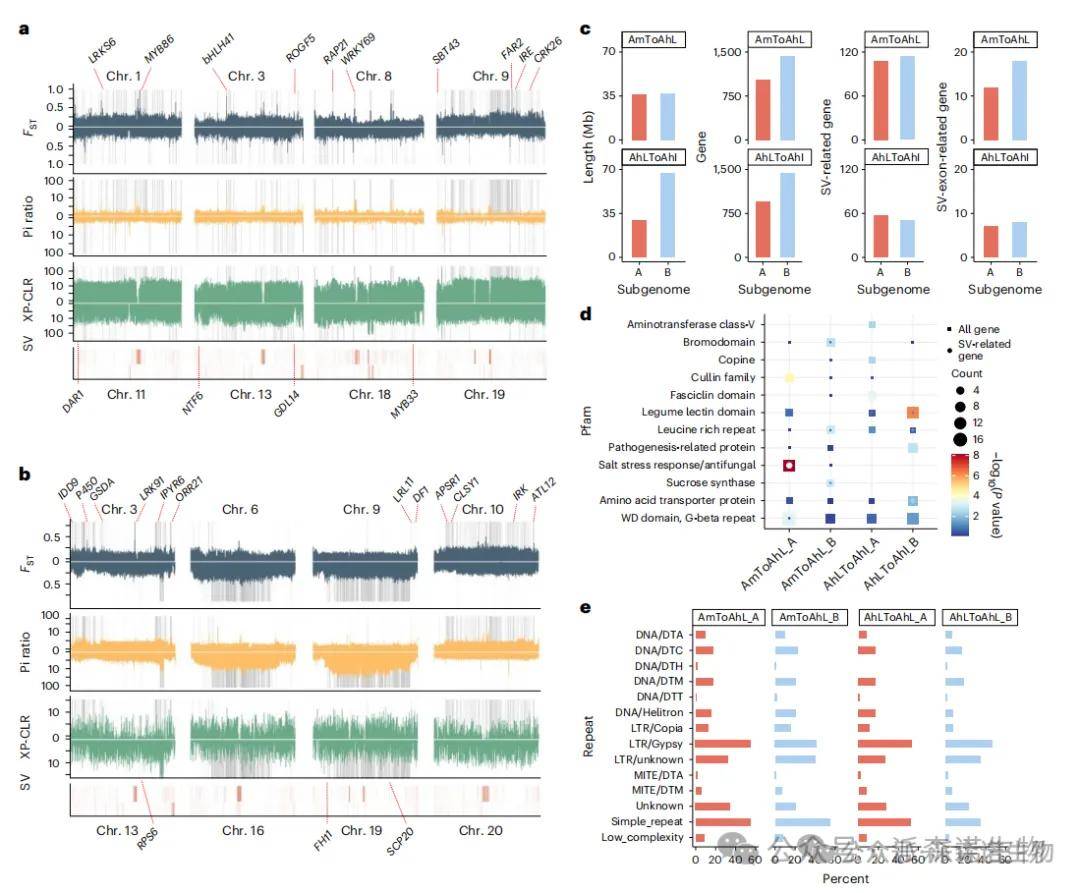
图4 驯化过程中全基因组的选择性清除
5、CKX6 基因中的结构变异降低其表达并促进种子膨大
研究鉴定出117个与种子重量显著相关的SV或InDel,其中染色体3上一个SV(P = 1.84 × 10⁻⁴)显著与种子重量关联。该SV附近的AhCKX6基因在大粒品种中3′-UTR存在两个特异插入序列,75%的地方品种和改良品种携带该插入,而在61个野生品种中完全缺失。表达分析显示,小粒品种中AhCKX6表达显著高于大粒品种,而大粒品种的细胞分裂素含量更高。功能推断表明,这些插入降低CKX6表达,从而减少细胞分裂素降解,提升其积累水平,进而加快细胞分裂,促进种子发育。
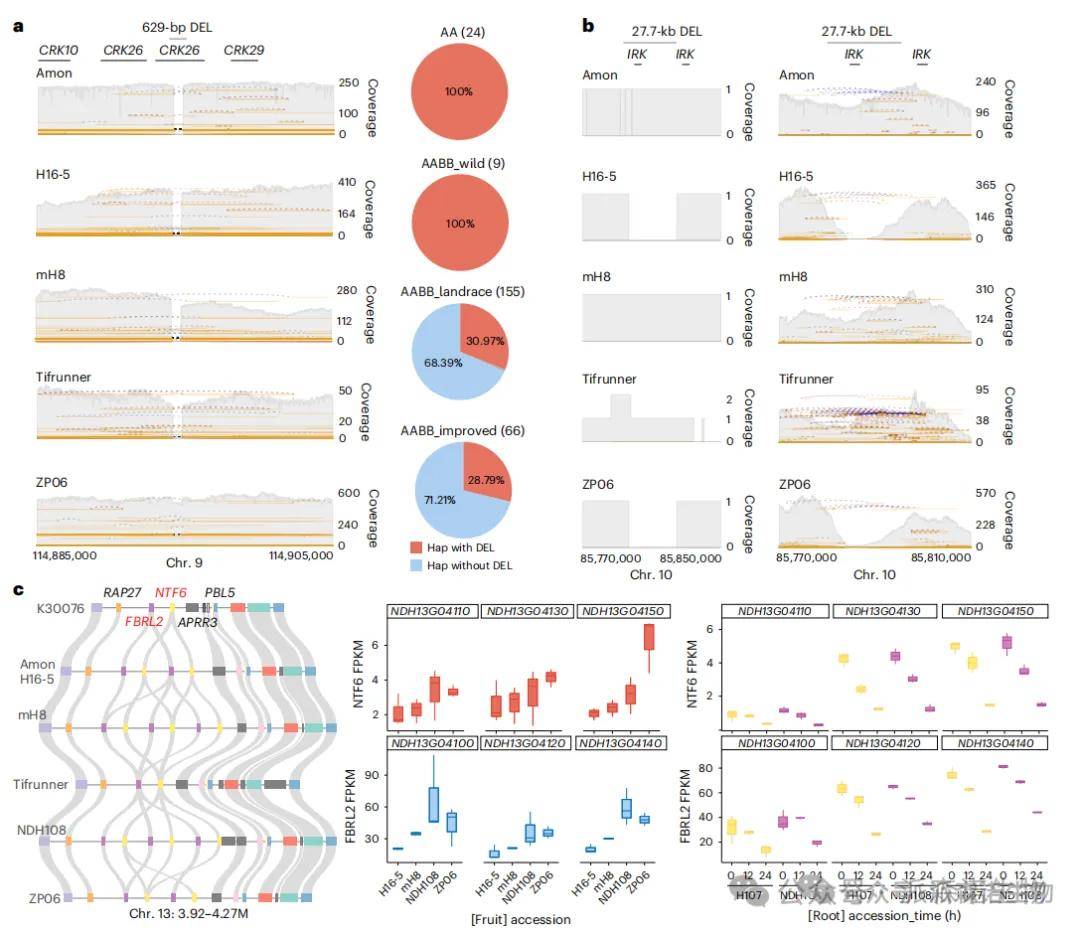
图5 选择清除区域中具有结构变异的性状相关基因
6、AhARF2-2 负调控种子大小
在SV-GWAS中,共鉴定出73个与种子大小相关的SV,其中NDH08G29450基因(AhARF2-2)中的一个外显子突变最显著(P = 3.63 × 10⁻⁶)。ZP06品种的HapII型在第12、13外显子中存在275 bp缺失和提前终止密码子,导致AUX/IAA结构域丢失,使蛋白失去与AhIAA13和AhTPL的互作能力。该突变显著影响种子长度和百粒重。表达分析显示,AhARF2-2在HapII中表达更高,但对下游GRF5的抑制较弱,GRF5表达在HapII中更高,从而促进种子膨大。转基因验证中,HapII型拟南芥种子长度和面积显著大于HapI型。综上,AhARF2-2通过招募TPL抑制GRF5表达,而突变型丧失该调控能力,从而解除抑制、促进种子发育。
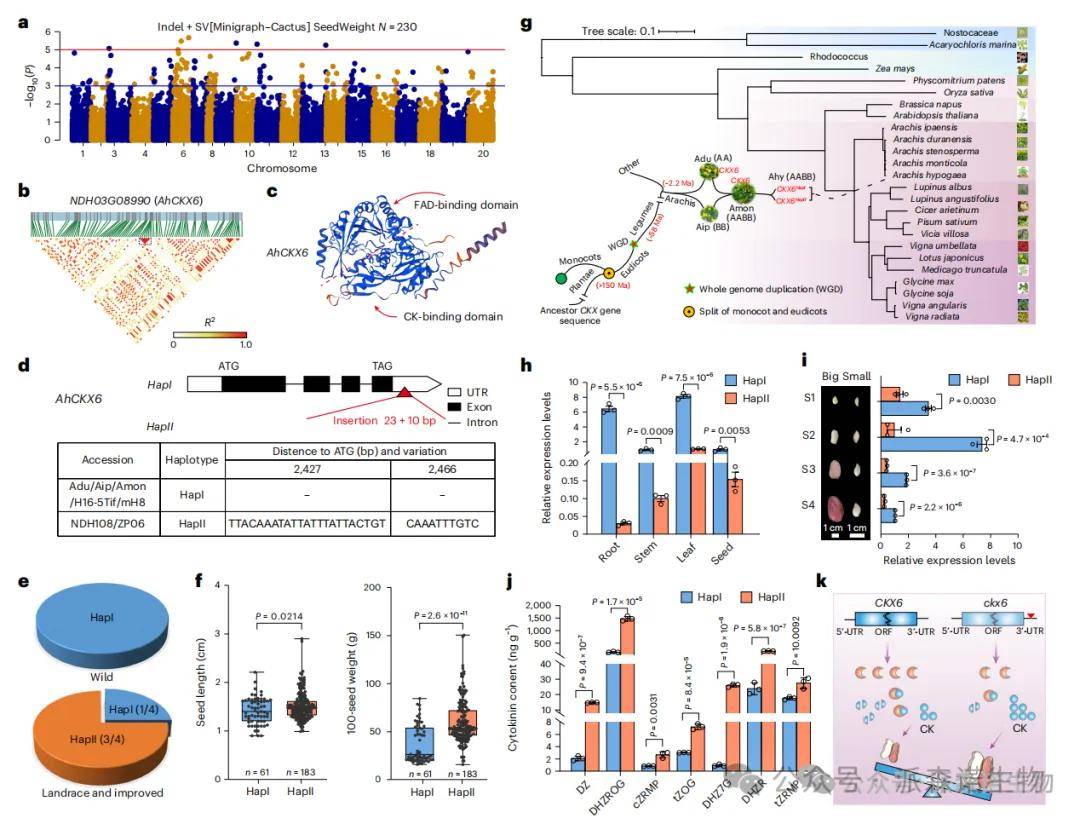
图6 结构变异全基因组关联分析
总结
研究人员基于泛基因组分析揭示了花生种子大小和重量相关的结构变异。研究利用8个高质量基因组和269份不同种子大小的重测序数据,鉴定出22,222个核心或软核心基因家族、22,232个分布基因家族和5,643个私有基因家族。研究发现A亚基因组中的结构变异(SVs)频率高于B亚基因组,并鉴定出1,335个与驯化相关的SVs和190个与种子大小或重量相关的SVs。特别地,AhARF2-2基因中一个275 bp的缺失导致其与AhIAA13和TOPLESS的相互作用丧失,减少了对AhGRF5的抑制作用,从而促进了种子的扩张。这一高质量的泛基因组为花生及其他豆类作物的遗传改良提供了重要的基础资源,并为理解花生的驯化机制和遗传多样性提供了新的视角。


















