




















文章信息
英文题目:Haplotype-resolved, gap-free genome assemblies provide insights into the divergence between Asian and European pears
中文题目:单倍型分离、无间隙的基因组组装为亚洲和欧洲梨的分化提供了见解
发表期刊: NATURE GENETICS
影响因子:29.0/Q1
发表时间:2025年8月
涉及组学:T2T基因组、群体进化、SV-GWAS、转录组。
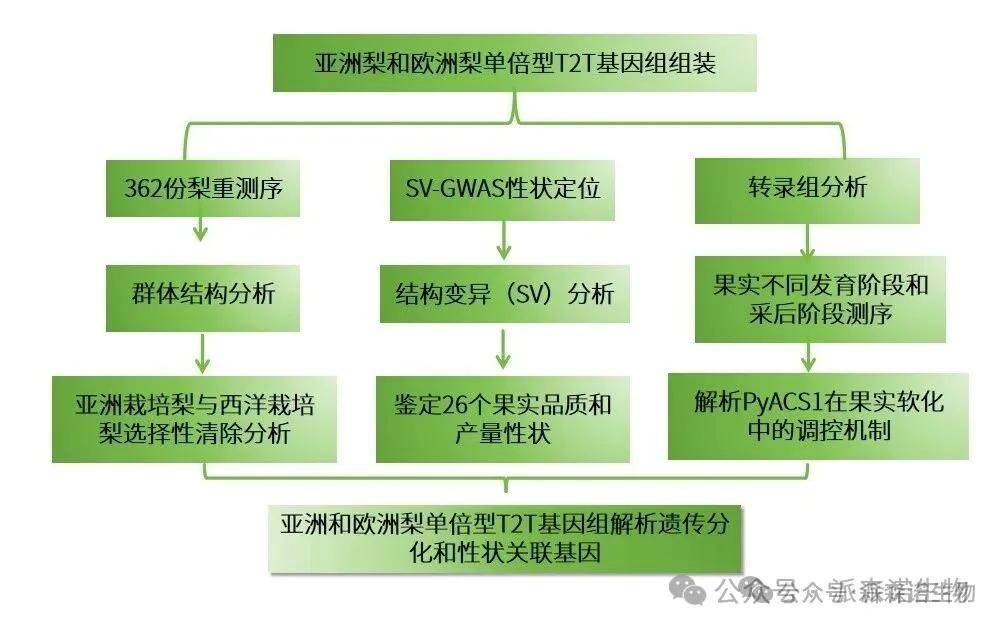
技术路线
摘 要
梨属植物(Pyrus spp.)是具有广泛遗传多样性的自交不亲和作物。由于高杂合度和技术限制,参考基因组中存在缺失区域。本研究展示了亚洲梨品种‘丹参梨’(Pyrus bretschneideri)和欧洲梨品种‘马克斯红巴特利特’(Pyrus communis)的端粒到端粒单倍型解析基因组。单倍型特异性基因在转座子含量、甲基化模式和表达水平方面与双等位基因存在显著差异。等位基因特异性表达分析表明,显性效应对梨果品质形成至关重要。对362个样本的群体分析显示,种间基因渗入显著增加了梨属植物的遗传多样性。我们构建了基于图谱的基因组,并鉴定出与农艺性状相关的结构变异。研究发现亚洲与欧洲梨在启动子区域存在286bp插入,以及PyACS1基因的差异表达,这两种梨具有不同的果肉软化特性。进一步实验证实了PyACS1在果肉软化中的作用。总体而言,本研究揭示了遗传变异的奥秘,为梨树改良提供了新思路。
前言
梨作为全球重要果树,具有自交不亲和性,可分为亚洲梨和欧洲梨两大类群,两者在果实形态、糖酸组成及采后软化等性状上差异显著。早期发布的“砀山酥梨”(2013)和“巴特莱特”(2014、2019)基因组仍存在缺口与覆盖不全的问题。2019年首次报道了亚洲梨的阶段二倍体组装,而近期基于马赛克策略完成的“砀山酥梨”与“康弗伦斯”梨的T2T基因组仍无法完全解析单倍型间差异。此外,梨的群体演化历史与系统发育关系仍有待深入探究。
本研究分别对亚洲梨品种“砀山酥梨”(DS)和欧洲梨品种“麦克斯红巴特利”(MRB)完成了单倍型分型的T2T高质量基因组组装,显著提升了基因组完整性与准确性。结合362份梨种质的重测序数据,我们系统揭示了梨的演化历史与种群动态,并构建了基于图谱的基因组,用于鉴定亚种间存在频率差异的结构变异及其与果实性状、基因表达的关联。研究发现,位于PyACS1启动子区的一段286 bp插入可能延缓欧洲梨成熟并促进软化。本研究为理解梨的杂种优势、演化机制及关键农艺性状的遗传基础提供了宝贵资源,并将助力梨育种实践。
研究内容
1.梨单倍型T2T组装
研究人员基于高杂合度的“砀山酥梨”(DS,1.23%)和西洋梨(MRB,1.33%),整合PacBio HiFi、ONT超长读长和Hi-C数据,利用hifiasm初步组装出二倍体单倍型,再通过Verkko和HiCanu填补缺口与端粒区域。所有填补区域均获ONT和HiFi读长均匀覆盖支持。最终每个单倍型均被组装为17条对应染色体的contig,基因组大小在497.50–505.55 Mb之间,contig N50 ≥ 28.96 Mb。相比已发表基因组中18.89–24.13 Mb未组装区域,本次组装在contig N50、比对率、BUSCO评分及LTR组装指数等方面均表现优异。通过端粒重复序列鉴定,DS和MRB分别检出34和33个端粒(图1a–b),标志着DS与MRB的T2T单倍型基因组已完成。
转座元件(TEs)在DS和MRB中占262.86–272.23 Mb,以LTR为主。共预测42,675–43,602个蛋白编码基因,BUSCO完整性达97.90%–98.60%,为研究着丝粒、S位点等多态区域提供基础(图1c–d)。在新组装区域中,DS与MRB分别鉴定出577/563和697/804个基因,多分布于高TE含量与密度区(77.33%–84.06%)。这些包括调控发育与应激的细胞分裂周期蛋白,及参与糖分解的UDP-葡萄糖类黄酮3-O-葡萄糖基转移酶。
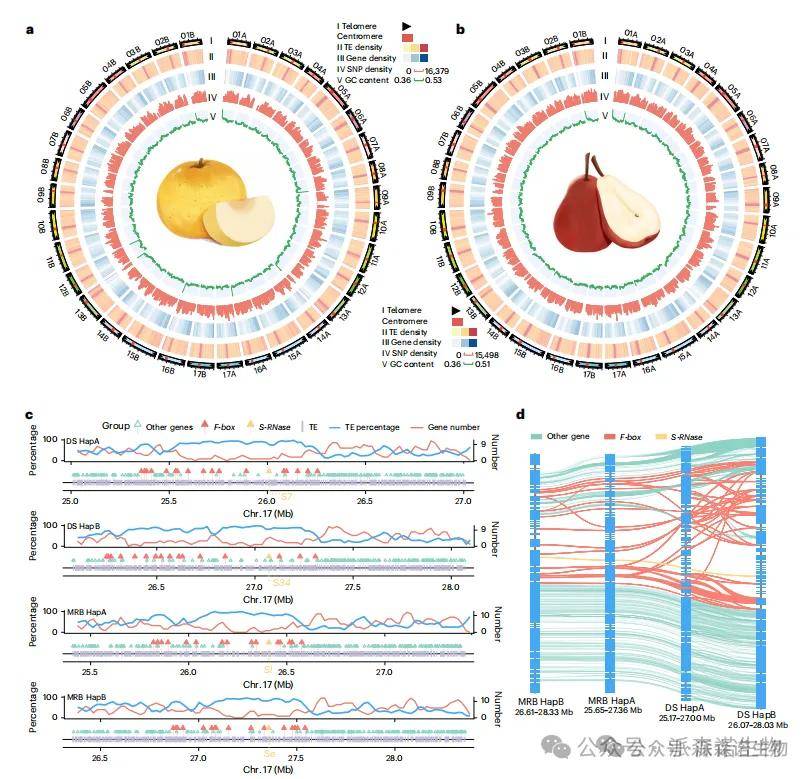
图1 DS和MRB基因组组装
2.鉴定单倍型特异性和ASE基因
长期的杂交历史使梨的两个单倍型间积累了广泛的遗传变异和等位基因多样性。研究人员基于直系同源基因对应关系,共鉴定出33,710–34,036个CDS完全不同的双等位基因(图2a,b),其中6,691–8,087个基因在同一单倍型内存在CDS完全相同的等位基因对。此外,还发现1,703–1,994个单倍型特异性基因。与双等位基因相比,这些基因在上下游5 kb及基因本体区域具有更高的转座子(TE)含量(图2c),表达水平普遍较低,且DNA甲基化程度更高,其基因本体及侧翼55 kb区域在叶片和果实中均呈现更高甲基化,显示其表达可能受甲基化抑制,与已有研究一致。KEGG富集分析表明,这些基因显著集中于“植物-病原互作”和“环境适应”通路,GO术语富集于“对生物刺激的响应”。
在DS和MRB八个果实发育阶段中,分别有19.64和26.21%的基因呈现等位基因特异性表达(ASE)(图2d)。ASE基因可分为三类:HapDom、新功能化配对等位基因,以及其他类型(图2e)。木质素合成关键基因Pbre1_15G769200(C3H)和Pbre1_13G264800(4CL)在果实早期(S1–S2)表达均显著高于另一单倍型中的等位基因(图2h)。
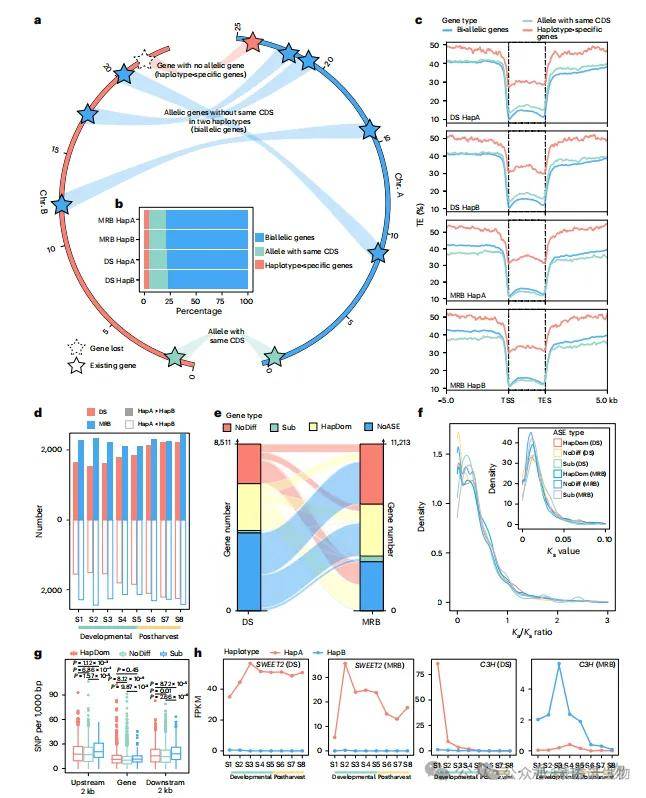
图2 | 田间试验验证的三种GS方法的优选增益。
3.亚洲梨与欧洲梨之间的基因组进化
为解析亚洲梨与欧洲梨的进化历史,本研究基于DS、MRB等12个物种的蛋白质编码基因进行了比较分析(图3a)。系统发育显示,DS与MRB约在360万年前分化,二者分别新增767和740个基因家族。DS中扩增的基因家族主要富集于“植物次生代谢物合成”“植物-病原互作”及“应激响应”等通路;MRB则显著富集于“果糖与甘露糖代谢”“碳水化合物代谢”及“次级代谢过程”。
两亚种间显著的进化分歧反映在基因组结构差异上(图3b)。基于4种单倍型构建的图谱基因组共鉴定出103,069个SV位点,其中57.42%位于基因及其调控区(±22 kb),61.27%与转座元件重叠(图3c)。研究还发现12个长度超过100 kb的大规模倒位,均经ONT或Hi-C验证(图3d)。特别值得注意的是,chr. 17上S位点区域存在一个0.58 Mb的倒位,该结构特异分布于DS的HapA和HapB之间,可能通过抑制重组维持自交不亲和相关基因(SFB与S-RNase)的连锁(图3d)。
基于362份梨种质的重测序数据,进一步对比到图谱基因组进行SV基因型鉴定,最终保留83,369个高质量SV位点(等位基因频率>0.05,缺失率<0.2)。以DS HapA为参考时,76.82%的SV在亚洲梨中出现频率超过0.5,而在欧洲梨中仅为42.73%(图3f)。共有35,481个SV在两类群体间呈现显著频率差异(FDR<<0.001,倍数变化>2)。
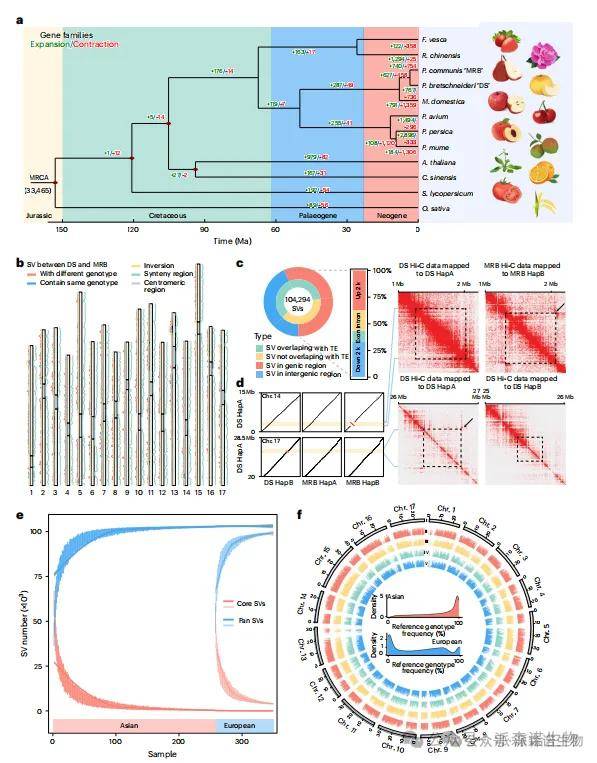
图3|DS和MRB之间的分歧和SVs。
4.进化历史与遗传渗入
为进一步探讨亚洲梨与欧洲梨之间的遗传渗入事件、种群动态历史及驯化过程,我们基于SNP和SV构建了系统发育树。结果显示,这362份梨品种被划分为两个明显不同的分支:分支I包含大部分亚洲梨,而分支II则由欧洲梨组成(图4a)。高固定度统计指标(FST)及主成分分析结果均支持亚洲梨与欧洲梨之间长期的分化历史(图4b)。值得注意的是,栽培乌苏里梨的核苷酸多样性(4.80 × 10⁻³)显著高于野生乌苏里梨(3.26 × 10⁻³)。此外,结构分析揭示了栽培梨的历史杂交事件(图4a),表明栽培乌苏里梨具有两种遗传背景,分别源自野生乌苏里梨和栽培砂梨(或北非梨)。Treemix分析进一步检测到北非梨向乌苏里梨的基因流,提示基因渗入事件可能在乌苏里梨的驯化过程中发挥了重要作用。
通过对种群动态历史的推断,我们发现亚洲梨(除野生乌苏里梨外)及欧洲梨的种群有效大小(Ne)经历了三次显著下降(图4c)。梨的驯化导致其野生祖先在糖酸含量、石细胞和果实大小等性状上发生显著变化。本研究在亚洲栽培梨(AC)与西洋栽培梨(AW)之间鉴定出21.31 Mb的选择性清除区域,在欧洲栽培梨(EC)与西洋栽培梨(EW)之间鉴定出14.59 Mb的区域(图4d,e),并在其中识别出123个与果实大小、发育、石细胞及糖酸代谢相关的候选基因。
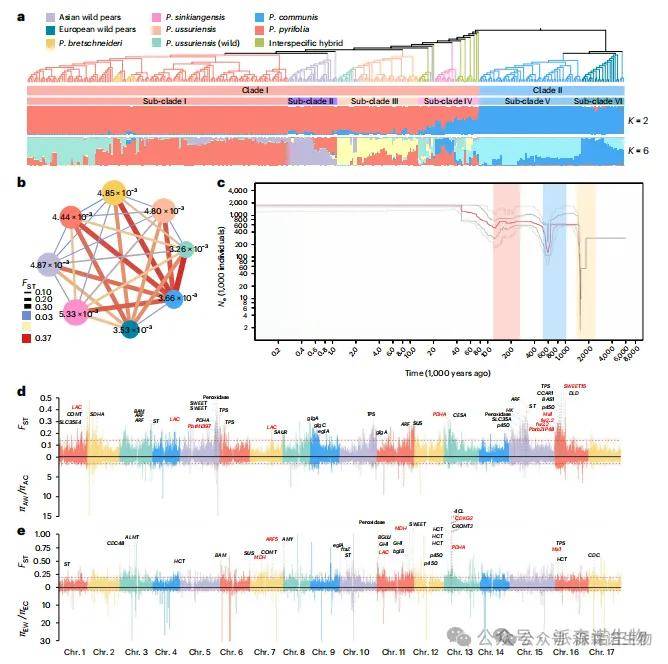
图4|不同梨类群体的系统发育关系、种群历史和选择性清除。
5.与农艺性状相关的SVs
本研究利用83,369个高质量SV,首次通过SV-GWAS揭示了与梨果实性状相关的结构变异。分析发现,第2号染色体上一个66 bp缺失与果实横径(FTD)显著相关(图5a)。该缺失位于AC梨的选择清除区内(图5b),在Ppy和Pbr中出现频率低于AW类群(图5c),且非缺失型品种的FTD显著更高(图5d)。其下游基因Py TPR编码一种四肽重复蛋白,已知参与植物发育及细胞伸长过程。该基因在非缺失型资源中表达水平显著更高(图5f),表明Py TPR可能在亚洲梨驯化中被选择,并影响果实发育与风味积累。
此外,一个1,016 bp缺失与可溶性固形物含量(SSC)显著相关(图5g),缺失型种质SSC更高(图5h)。该区域内的基因PybZIP6(Pbre1_15G533000)属bZIP转录因子,已知调控梨SSC积累。PybZIP6在果实发育后期(S3–S5)高表达(图5j,补充图32),且该缺失在Pco中频率高于EW(图5k)。上述SV为梨果实品质改良提供了新的分子标记。
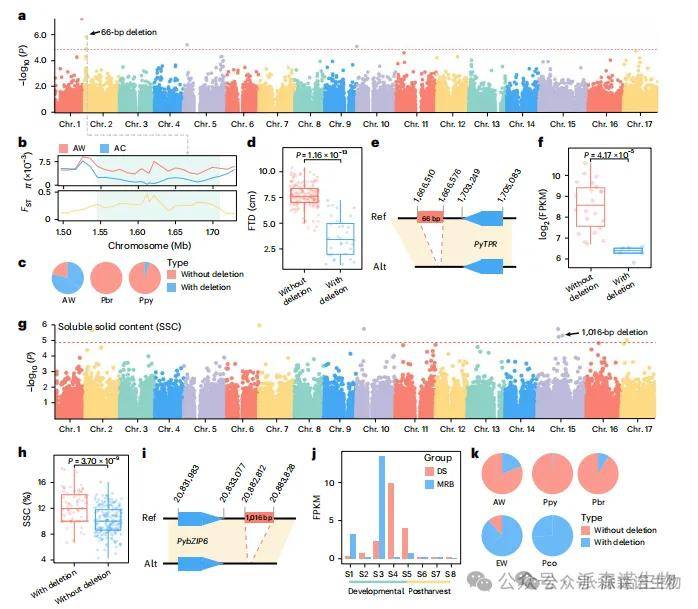
图5|FTD和SSC的SV GWAS
6.PyACS 1在果实成熟和软化中的作用
结构变异(SV)可能通过干扰调控区或基因区域影响基因表达。基于110份梨样本(62亚洲梨、38欧洲梨)的RNA-seq数据,研究发现上游2 kb区域的4,040个SV和外显子区的568个SV与基因表达显著相关(图6a);其中分别有70.79%和65.49%的SV在亚欧群体间频率差异显著(FDR<<0.001,FC>2),表明SV可能调控群体间基因表达。与亚洲梨不同,呼吸跃变型的欧洲梨依赖乙烯启动成熟和软化。研究鉴定出5个携带SV的乙烯合成相关基因,包括编码限速酶ACS的PyACS1(Pbre1_15G535800)。该基因在欧洲梨中表达显著高于亚洲梨(图6b),种间杂交后代表达更高,提示超显性表达。PyACS1在MRB果实后期(S4–S5)和采后(S6–S8)高表达,在DS各阶段均低表达(FPKM<0.1;图6c)。过表达PyACS1的番茄果实成熟更快(图6d,e)、乙烯释放增多(图6f)、硬度下降(图6g),证实其促进成熟与软化。
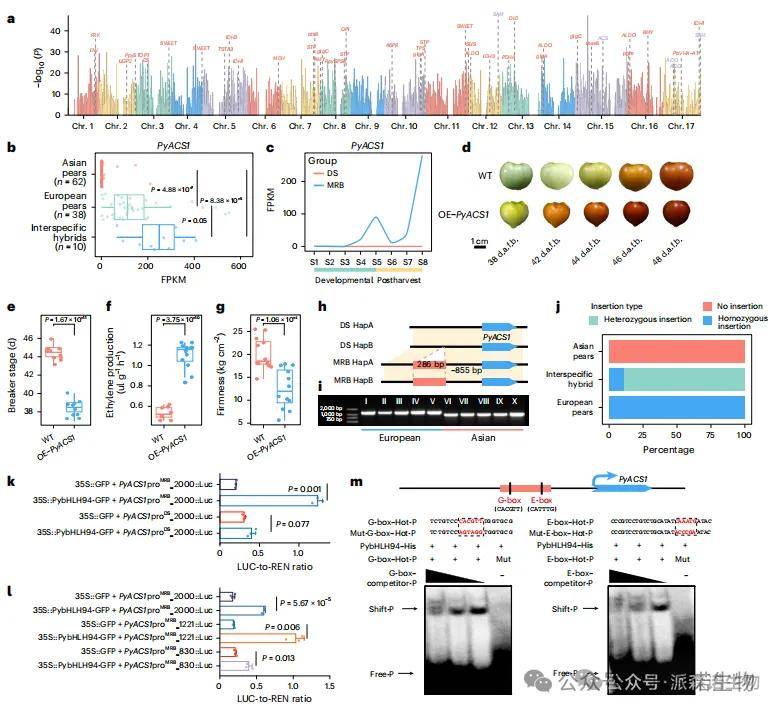
图6|与基因表达相关的SVs对欧洲梨果实成熟和软化的功能影响
总 结
本研究构建了两个栽培品种“Dangshansuli”和“Max Red Bartlett”的高保真组装全基因组序列,并利用重测序数据鉴定了362个不同品种间的结构变异,揭示了遗传多样性来源。同时,通过基因组比较发现了这两个品种之间存在显著差异,如染色体倒位、插入和缺失等。此外,通过对基因表达谱的分析,发现了一些与果实成熟和软化相关的基因,并验证了其中一个关键基因PyACS1在果实成熟和软化过程中的重要作用。最后,还发现了一个286bp的插入序列,该序列存在于所有欧洲梨及种间杂交种质中,但在亚洲梨中则未检测到,且该序列的存在可以被一个特定的转录因子PybHLH94所调节。总之,本研究为深入了解梨的遗传多样性及其进化提供了重要的基础数据。
参考文献:Sun M, Cao B, Li K, et al. Haplotype-resolved, gap-free genome assemblies provide insights into the divergence between Asian and European pears. Nat Genet. 2025 Aug;57(8):2040-2051. doi: 10.1038/s41588-025-02273-4.
