

2017-12-06
越来越多的研究成果表明非编码RNA(lncRNAs)和环状RNA(circRNAs)参与胚细胞的发育。但是鲜少有研究lncRNAs和circRNAs在胚胎生殖细胞的自我更新和细胞分化中的功能和机制。因此,本文通过高通量测序,研究mRNAs,lncRNAs和circRNAs在雄性和雌性小鼠胚胎生殖细胞中的表达分析。在胚胎生殖细胞中鉴定了18,573个新lncRNAs和18,822个circRNA,并通过RT-PCR手段验证了这些lncRNAs和circRNAs的存在。结果表明雄性和雌性小鼠的胚胎生殖细胞有着相似的GDNF(神经胶质细胞源性的神经营养因子)信号传导机制。随后发现8,115个mRNAs,3,996个lncRNAs和921个circRNAs表现有性别偏向的表达方式,这可能和生殖细胞获得独特性别属性需要分化成配子有关系。基因本体论(GO)和KEGG通路富集分析揭示了这些性别偏向的lncRNAs和circRNAs的不同功能。之后利用生物信息构建了关联表达网络图,包括编码-非编码的共表达竞争内源性RNAs。共表达分析表明数百的lncRNAs和老鼠的胚胎生殖细胞的性别差异有关,包括lncRNA Gm11851,lncRNA Gm12840,lncRNA 4930405022Rik,和lncRNAAtp 10d。ceRNA网络图推测lncRNA Meg3和circRNA Igf1r会竞争性结合miRNA-15a-5p,增加靶基因Inha,Acsl3,Kif21b和Igfbp2的表达。这些发现为lncRNAs和circRNAs的研究提供了新视角,也为以后对胚胎生殖细胞中lncRNAs和circRNAs的调控机制的研究奠定了基础。
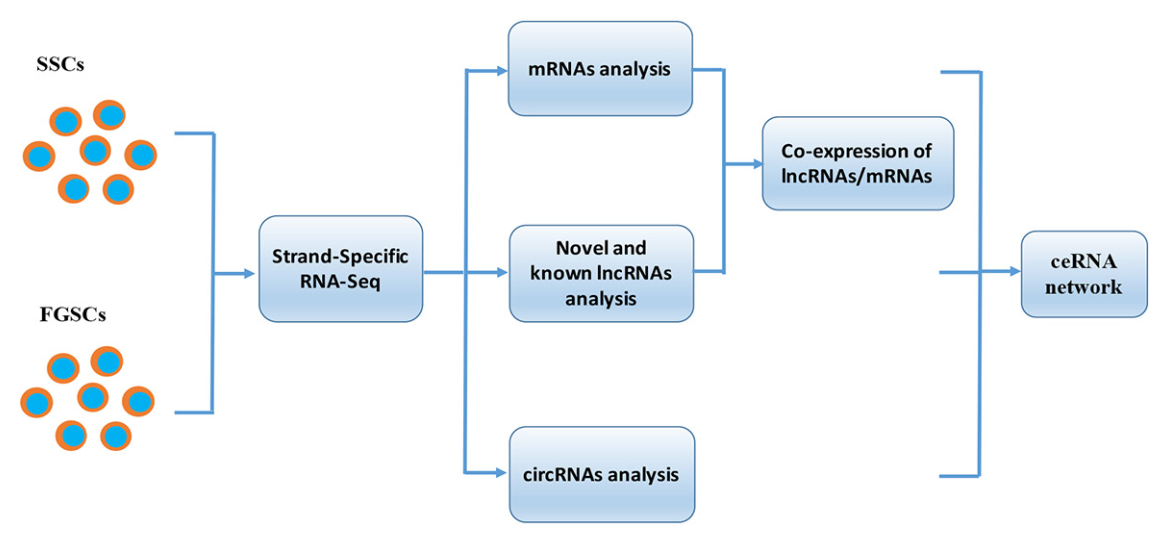
使用荧光活化细胞分选法分离SSCs和FGSCs。(图1A-1D)评估胚胎生殖细胞的方法如下:首先通过RT-PCR检测SSCs和FGSCs中的Mvh,Dazl,Fragilis,Otc4,Stella和Blimp1的基因表达。结果表明细胞中表达Mvh,Dazl,Fragilis,Otc4,Stella和Blimp1。然后,免疫荧光分析验证Mvh,Otc4和Dazl的表达。随后,基于前面的结果,检测18个生殖细胞基因表达,发现18个生殖细胞基因在SSCs和FGSCs中都有表达。最后,结合EdU的荧光免疫分析证明SSCs和FGSCs拥有繁殖能力,分离得到的SSCs和FGSCs中真正的胚胎生殖细胞占有相似的比例(>90%)(图1E-1J)。通过Illumina(双端)测序技术,分析两种胚胎生殖细胞中mRNAs和非编码RNAs的表达。SSC和FGSC文库分别产生了249,912,216和216,924,516条raw reads,GC含量分别是53.5%和50.7%。过滤之后,得到233,978,622(93.6%)和246,157,442(94.0%)条clean reads,用以做后续分析。SSC和FGSC文库比对上小鼠参考基因组(UCSC mm10)的比例分别是86.7%和91.1%,利用Scrupture和Cufflinks组装了58,364个转录本。
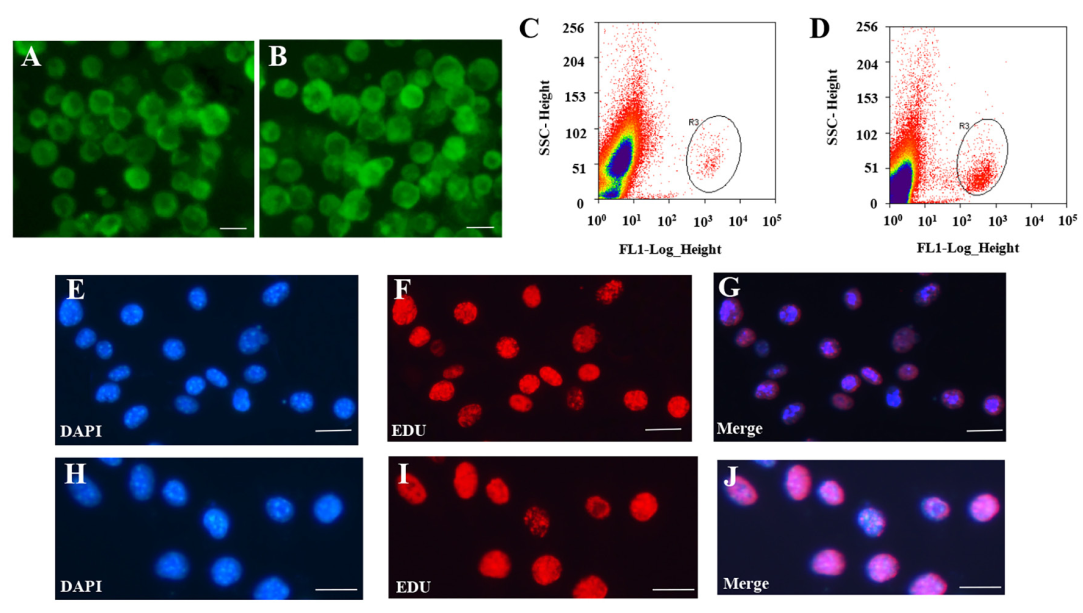
图1 FGSCs和SSCs的分离和纯化
将6个样本的数据在RefSeq,Ensembl,或者NoncodelncRNA数据库中搜索以确认新lncRNA。为了将58,364个转录本中lncRNA的假阳性最小化,使用严格的过滤方法去掉没有lncRNA所有特性的转录本。去除长度小于200bp,只有一个外显子,少于3次覆盖率的转录本,使用CPC和CNCI软件对剩余的转录本进行编码潜能预测,鉴定出18,573个新lncRNAs和5803个已知lncRNAs(图2A)。新lncRNA的平均长度是1,427个核苷酸,和已知lncRNAs长度相似(图2B)。lncRNAs和mRNAs的开放阅读框(ORF)的平均大小分别是86.24bp和394.84bp,表明mRNA的ORFs要明显长于lncRNA的ORF。LncRNA不能编码蛋白质因为缺少重要的ORFs。在18,573个新lncRNAs中,我们发现4300个新lncRNAs表达量FPKM值>1,12,022个新lncRNAs表达量FPKM值>0.5。随机挑选SSCs和FGSCs文库中鉴定的6个新lncRNA转录本,通过RT-PCR验证。图2C中,6个lncRNA转录本进行扩增获得预期大小的转录本。另外用RT-PCR扩增TCONS_00083536,通过RACE技术克隆基因序列,获得了1163bp长度的序列。克隆得到的序列能和RNA-seq数据库完全匹配。qRT-PCR结果表明两组样本中选择的lncRNAs的表达模式和lncRNAs的FPKM值是一致的,测序结果合qRT-PCR结果是相关的(图2D)。以上分析结果表明我们筛选lncRNA的方法是严谨的,大部分的lncRNA是在体内真正表达的。
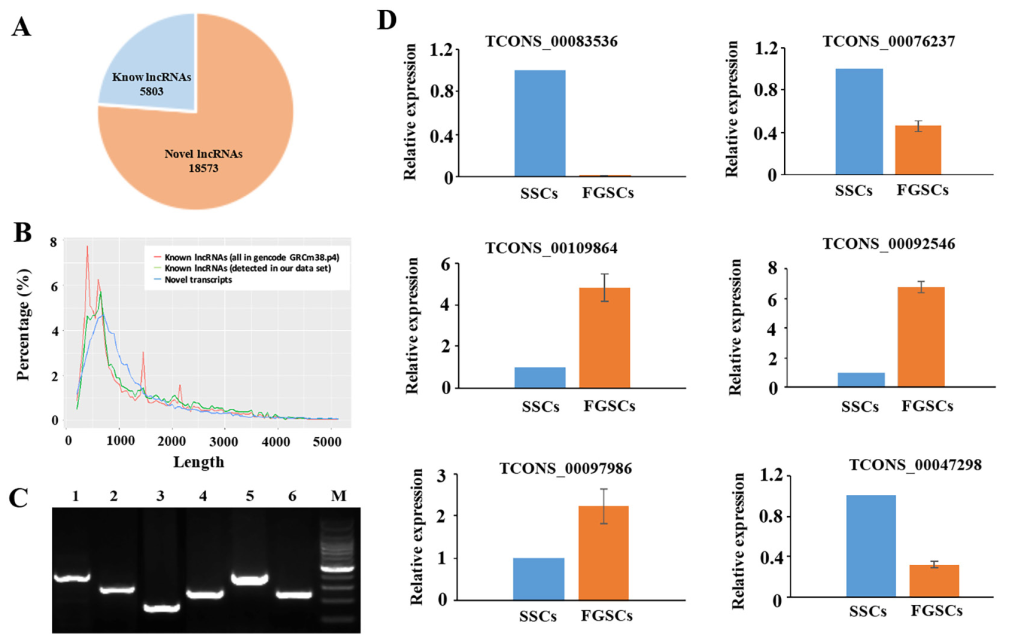
图2 新lncRNA的鉴定和验证
LncRNA和mRNA转录本分布在所有的小鼠染色体上。分析结果表明新lncRNAs在所有染色体上广泛分布,每条染色体上lncRNA的表达比率要比mRNA的表达比率高很多(图3A)。这些分析结果证明在雄性和雌性小鼠胚胎生殖细胞中lncRNAs基因的转录与mRNA基因的转录一致性很好。依据lncRNA相对于最近的蛋白编码基因的位置,将lncRNA分类:正向重合lncRNA,双向lncRNA,反向lncRNA和基因区间lncRNA。测序得到的新lncRNAs中,主要是基因间区lncRNAs ,不同类型的lncRNAs在SSCs和FGSCs中数量是相似的(图3B)。
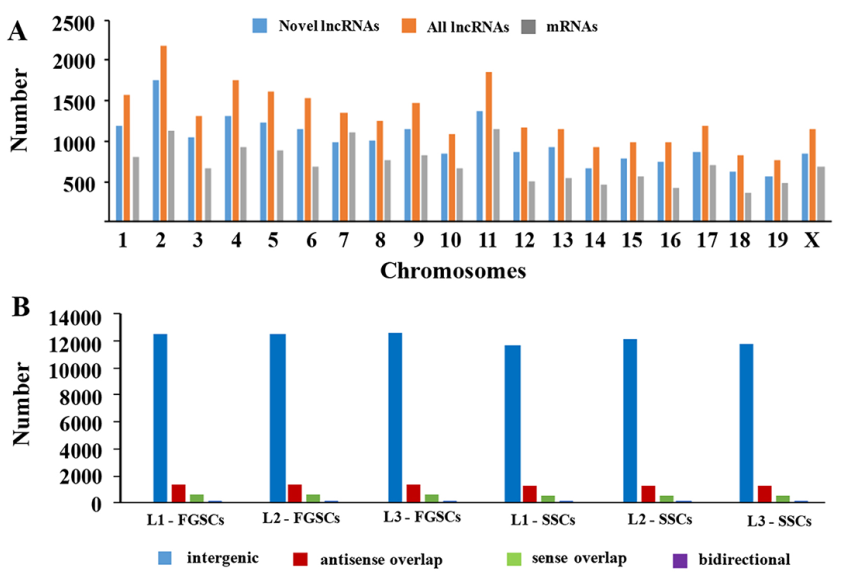
图3 新lncRNA染色体分布和分类
前期的芯片数据表明SSCs和FGSCs中所有的基因表达都是相似的。高通量测序数据验证了这个结论。所有的基因表达模式分析表明在SSCs和FGSCs中有些基因的mRNA和lncRNA水平是有相似的表达模式(图4A,4B)。对高度共表达的mRNAs和lncRNAs进行功能富集分析,发现主要富集在细胞周期,细胞增殖和细胞分裂相关的GO term上,意味着SSCs和FGSCs有着相似的胚胎生殖细胞维护机制,这和之前的芯片数据分析结果一致。
GDNF是促进SSC进行自我更新的第一优先级的外在因素,主要通过PI3K-Akt,Ras/ERK1/2和SFK通路促进SSC自我更新。为了验证FGSCs是否和SSCs有相同的GDNF信号机制,在7天中去除培养基里面的GDNF,发现结果和SSCs类似。去除GDNF一个星期导致细胞数量严重减少(1.0× 105减少到0.4× 105)(图4C-4E)。随后,将GFRα1(GDNF家族受体alpha 1)通过免疫荧光染色观察其在FGSCs表面的表达(图4F)。之前有研究表明对GDNF响应最明显的基因有Bc16b,Lhx1,Etv5和Egr3。基于以上信息,进一步发现去除GDNF一星期引起FGSCs中这些也和自我更新相关的基因表达水平减少。这些结果说明FGSCs和SSCs有着类似的GDNF信号传导机制。
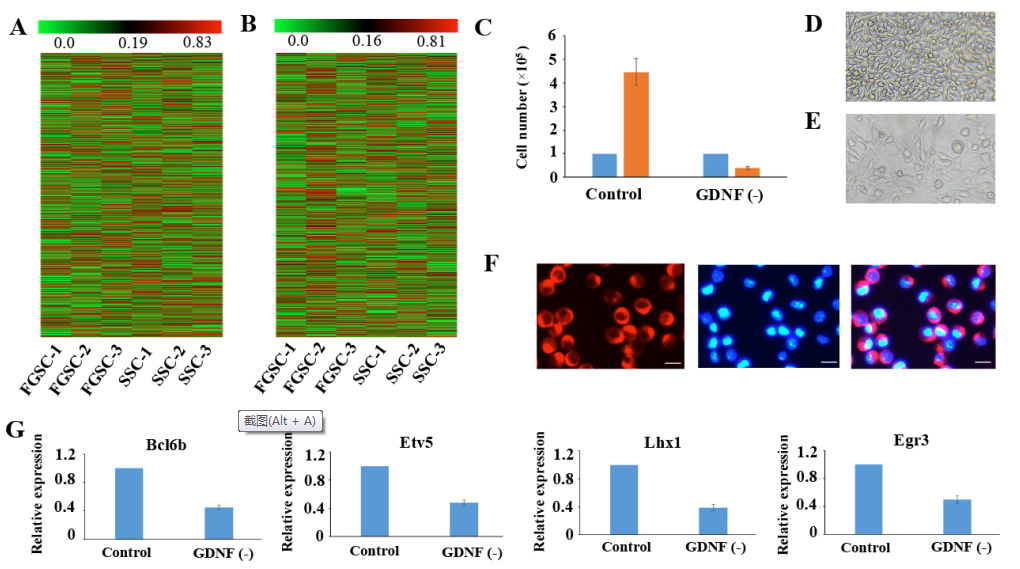
图4 SSCs和FGSCs中mRNA和lncRNA的表达分析及共有的自我更新机制研究
链特异性高通量测序提供精准的基因表达分析,这有利于挑选在雄性和雌性胚胎生殖细胞中有性别偏向性表达的基因,以做特殊性性别表达分析。通过lncRNA和mRNA分析,发现有8,115个mRNAs和3,996个lncRNAs(包含3,695个新lncRNA)表现出性别偏向性表达。其中,1,500个lncRNAs(包含1,364个新lncRNAs)和4,221个mRNAs表现出雄性偏向性表达,2,496个lncRNAs(包含2,331个新lncRNAs)和3,894个mRNA表现出雌性偏向性表达(图5A,5B)。雄性偏向性表达的lncRNAs优势比是1.22(雄性偏向性表达的RNAs/非雄性偏向性表达的RNAs),说明雄性偏向性lncRNAs在常染色体中富集。mRNA也表现出类似的趋势。SSC和FGSC比较发现雌性偏向性lncRNAs和mRNAs在X染色体上显著过表达。
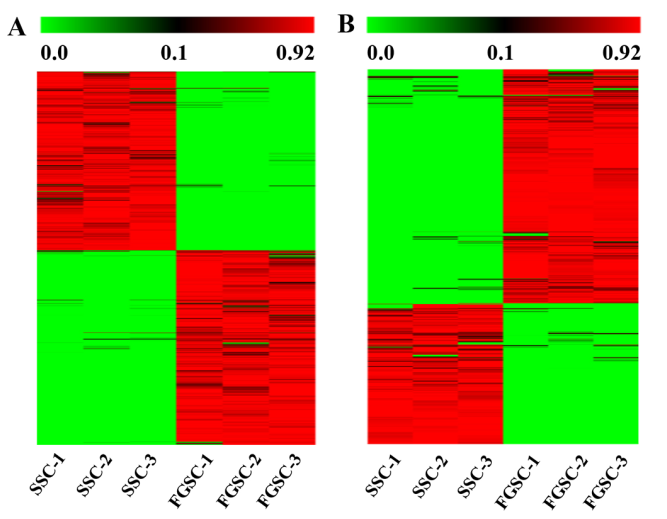
图5 转录组学分析性别偏向性表达的mRNAs(A)和lncRNAs(B)的表达丰度
LncRNAs通常和关联的mRNA同时转录组,可以调节重合或者邻近的mRNAs的转录组。所以,lncRNAs的功能可以通过关联的mRNAs顺式调控或者反式调控作用反应出来。基于GO和KEGG通路注释靶基因,进行预测性别偏向性表达的lncRNA功能分析。在GO分析中,lncRNAs最频繁的预测功能主要是和行为,生物粘附,生物阶段,和生物调节有关。功能聚类图见图6A。性别偏向表达的mRNAs和lncRNAs富集在和遗传印记及遗传印记的调控有关GO terms中,这表明遗传印记的差异可能调控SSCs和FGSCs中性别偏向基因的表达。
和FGSC相比,SSC中总共有3,996个性别偏向表达的lncRNAs比对到204个KEGG通路(图6B)。鉴定到的KEGG通路涉及糖代谢,蛋白代谢和脂质代谢通路,也包含类固醇合成,甘油磷脂代谢,脂肪酸代谢,三羧酸循环,甘油酯代谢,维生素A代谢,戊糖和葡萄糖醛酸转换。SSCs和FGSCs主要差异是胚胎细胞分化需要的性别特异属性;SSCs分化成精子,FGSCs分化成卵母细胞。结果提议糖代谢,蛋白质代谢和脂质代谢通路在细胞分化过程中占很大的比例,lncRNAs可能会通过这些信号通路影响性被特异属性。
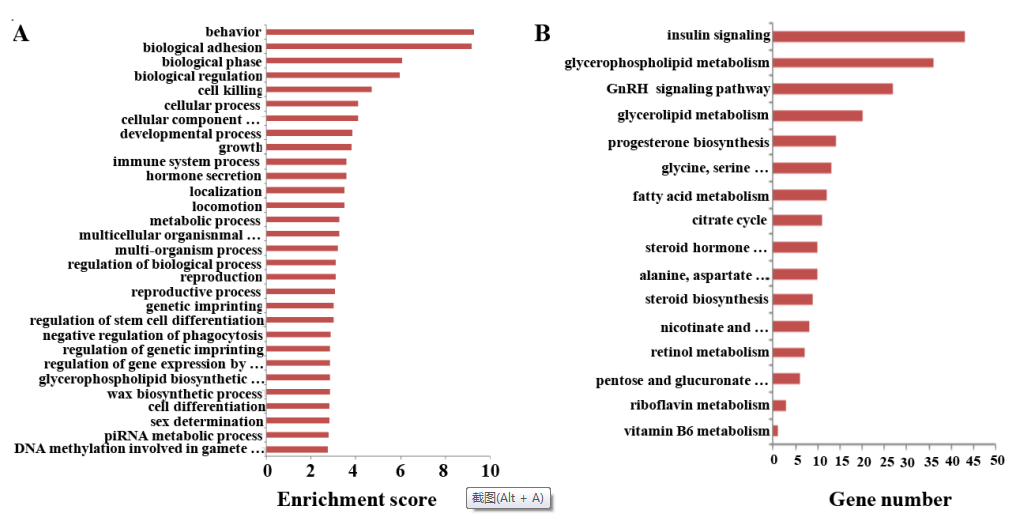
图6 性别偏向性lncRNAs的GO和KEGG通路分析
基于性别偏向表达的lncRNAs和mRNAs的关联分析,构建编码-非编码基因共表达网络图(CNC网络图)。选择前10个意义重大的性别偏向表达的编码基因建立CNC网络图(图7)。这些mRNAs涉及很多的生物过程,包括生殖细胞分化的增殖,调控,遗传印记调控,性分化和性染色质。网络图中可以看出上调的lncRNA Gm11851和参与遗传印记过程的Eed,Ndn和Peg3负相关,而lncRNA Gm12840是正相关。这说明一个mRNA会和1-10个lncRNAs有关联。
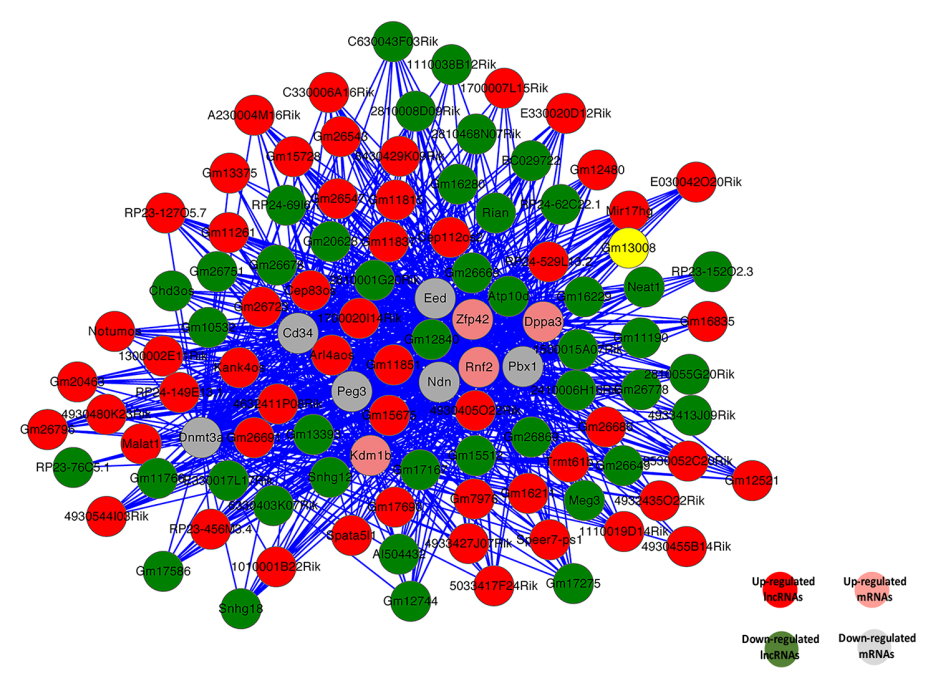
图7 10个重要的mRNAs和关联的lncRNAs的共表达网络图
使用CIRI分析RNA-seq reads,分析胚胎生殖细胞中circRNA。小鼠胚胎生殖细胞中从5,334个来源基因中衍生出18,822个circRNA。其中大部分是外显子circRNAs,只有345个是内含子circRNAs。研究发现9,812(52.13%)个circRNAs来源于同义链,9,010(47.87%)个circRNAs来源于反义链。之前的研究表明circRNAs通常缺少来源基因的第一个和最后一个外显子,在本研究中同样发现类似的结论:18,822个circRNAs中有18,803(99.9%)个circRNAs缺少对应来源基因的第一个或者最后一个外显子(图8A)。
为了验证circRNA的真实性,随机引物反转cDNA,设计多对外向引物,放大中cDNA锁尾插接外显子连接。通过RT-PCR,每对引物形成符合期望大小的单一条带,表示在SSC和FGSC样本中存在环状连接(图8B)。如果用oligo(dT)引物反转cDNA,得到的是只有poly-A的RNAs(图8B),不能得到circRNA。这说明circRNA没有poly-A尾。
将SSCs和FGSCs中circRNA来源基因的表达量和其他基因比较,发现circRNA来源基因的平均表达水平要显著高于非环状转录本的基因(图8C)。为了预测胚胎生殖细胞中这些circRNA来源基因的功能,进行GO和KEGG通路分析。结果表明circRNA来源基因在细胞中特异性表达。circRNA来源基因大多数的GO terms和KEGG通路主要是和胚胎生殖细胞的自我更新和分化有关,例如增殖,遗传打印,生殖细胞维持,细胞分化等。
CircRNA的表达分析表明921个circRNA表现出性别偏向性,其中245个表现出雄性偏向表达和676个雌性偏向性表达(图8D)。对有性别偏向性表达的circRNA来源基因做GO和KEGG通路分析,发现性别偏向性表达的circRNAs有不同的功能,这和分化成生殖细胞要求有性别特异属性有关(图8E和8F)。图8E展示的是排名前10的GO terms,图8F展示的是排名前10的KEGG通路。
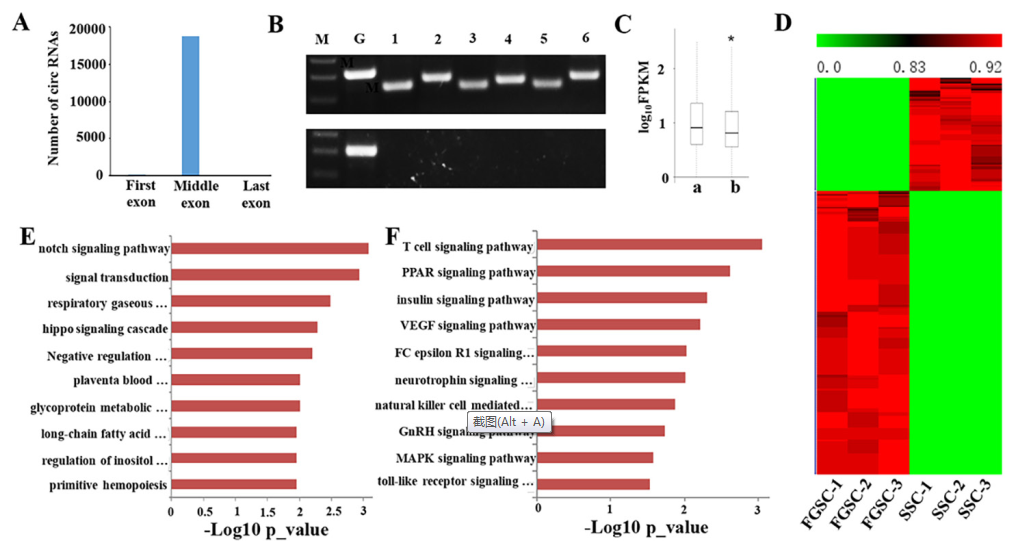
图8 胚胎生殖细胞中circRNAs的鉴定和功能分析
最近的研究表明RNAs利用miRNA应答元件(MREs)互相调控,这一机制叫做竞争内源性RNA(ceRNA)。通过整合高通量测序数据中的mRNAs,lncRNAs,circRNAs和miRNAs的表达分析和调控关系,构建ceRNA网络图(图9)。挑选了性别偏向性表达的60个lncRNAs和29个circRNA,共有一个MRE结合位点。例如,lncRNA Meg3和circRNAIgflr预测共同竞争miRNA miR-15a-5p,其靶基因是mRNA Inha,Acsl3,Kif21b和Igfbp2。包含有mRNAs,miRNAs,lncRNAs和circRNAs的ceRNA调控网络图,可能是胚胎生殖细胞获得分化为配子的性别特异性属性的重要节点。
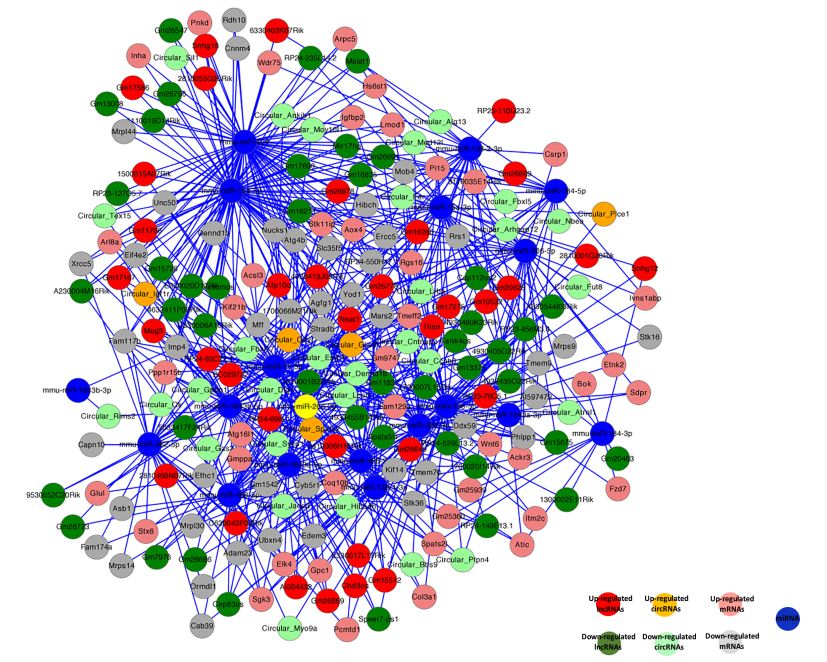
图9 胚胎生殖细胞中竞争内源性RNA网络图
文献来源:
Li X, Ao J, Wu J. Systematic identification and comparison of expressed profiles of lncRNAs and circRNAs with associated co-expression and ceRNA networks in mouse germline stem cells[J]. Oncotarget, 2017, 8(16):26573.


















