

2016-09-13
军团菌是一种致病菌,由于基因易位等因素具有约300个毒性蛋白。人们对其效应蛋白(effector)的认识较少。特拉维夫大学和芝加哥大学的研究者通过高通量测序得到了 38 株军团菌基因组的框架图,并对其编码效应蛋白的基因进行分析。研究者在 38 个菌株中总共预测得到 5,885 个编码效应蛋白的基因。进一步研究发现,38 个菌株共有的“核心”效应蛋白仅为 7 个,而其它大部分的效应蛋白具有种间特异性。同时,检测到很多保守的效应蛋白结构域和相互融合导致的未确定功能的效应蛋白。相关的研究成果于近期发表在Nature Genetics杂志上。
测序、进化和基因组描述
研究者首先对来自军团菌属的 41 个菌株进行全基因组分析(其中, 38 株菌为本研究中测序的菌株,3 株菌为已经测序的参考基因组菌株)。结果显示,这 41 株菌的基因组大小在 2.37 Mb~4.82 Mb,GC含量在 36.7%~51.1%。进化分析显示,这 41 株菌主要可以分为 3 个大的分支,见图1。
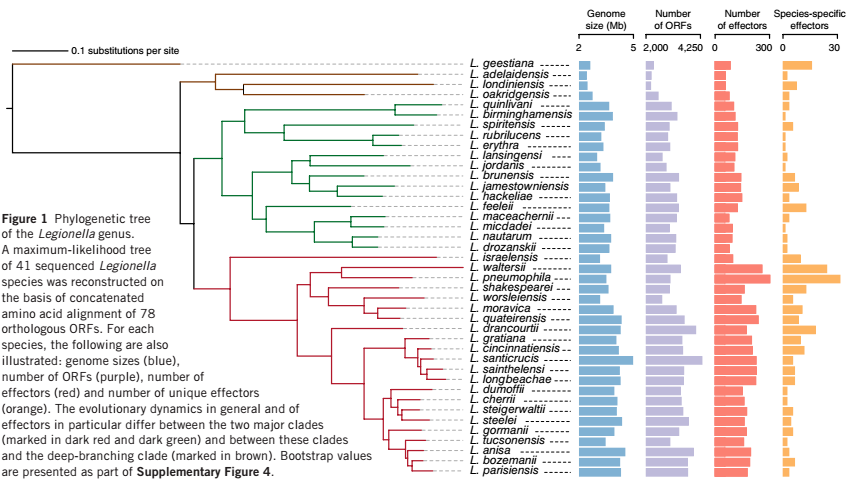
图1 军团菌属进化分析
Icm/Dot 分泌系统
Icm/Dot型分泌系统(IV B型分泌系统)是军团菌属主要的致病系统,由25个基因编码,分布于两个独立的基因组区域。对这两个区域的基因结构进行分析,发现Icm/Dot区域基因的顺序和方向高度保守。此外,在Icm/Dot 分泌系统中,发现了一些与该系统无关的片段,这些插入片段在不同菌株中的大小不等,见图 2。
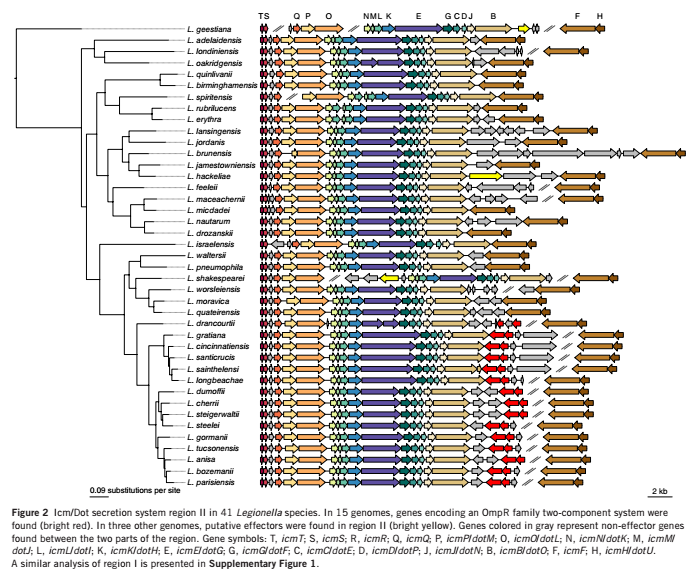
图2 Icm/Dot 分泌系统区域II 在 41 个军团属中的分布
军团菌属效应蛋白库
根据文献报道,军团菌属Icm/Dot 型分泌系统能转移约 300 种效应蛋白。采用机器学习方法对 41 个菌株进行效应蛋白预测,总共预测得到 5,885 个效应蛋白,可划分为 608 个军团属效应蛋白直系同源群(LEOGs)。其中,63% 的效应蛋白(269 个 LEOGs,包含3,715 个效应蛋白)为经过实验验证的效应蛋白(其功能直接采用实验验证或其直系同源蛋白采用实验验证);剩余的效应蛋白(339 个LEOGs,包含 2,170 个效应蛋白),代表新的效应蛋白,具有潜在的新的功能。在所有预测的效应蛋白中,只有 7 个效应蛋白在所有分析菌株中共有,称之为“核心效应蛋白”,见图 3。
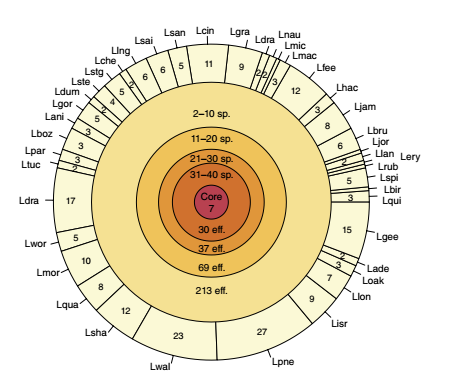
图3 军团菌属效应蛋白的 Venn 图
军团菌属 7 个核心效应蛋白
尽管在军团菌属中发现了 608 个 LEOGs,令人惊讶的是,只有其中 7 个为核心效应蛋白,这 7 个蛋白的进化历史和其物种树的进化历史一致。其中 1 个核心效应蛋白(LOG_00212,LegA3)能在目前所有已知的包含 Icm/Dot 分泌系统的细菌基因组中找到同源基因。另外 1 个核心效应蛋白(LOG_00334,MavN)能在军团菌目中的蟋蟀立克次小体菌(Rickettsiella grylli)基因组中找到同源基因,而不能在贝纳特氏立克次体菌(Coxiella burnetii)基因组中找到同源基因。其余的 5 个核心效应蛋白均不能在上述细菌基因组中找到同源基因,但是能在军团菌目之外的细菌基因组中找到同源基因。
为了进一步对这 7 个核心效应蛋白的功能进行分析,研究者在L. pneumophila 菌株中分别构建了这 7 个基因的敲除菌株(单个基因敲除),并研究这些敲除菌株在A. castellanii的细胞内生长情况。结果显示, LegA3 和 MavN 蛋白部分与该菌在该宿主的细胞内生长所必须。尽管这 7 个效应蛋白的准确功能未知,考虑到这 7 个效应蛋白在军团菌属进化过程中高度保守,推测它们可能在侵染过程中起关键作用。基因敲除实验发现,仅有 2 个核心效应蛋白与该菌在宿主细胞内的生长相关,这可能暗示,其余 5 个核心效应蛋白至少在L. pneumophila菌在A. castellanii细胞内的生长是冗余的。当然,这些核心效应蛋白可能在该菌在其他宿主细胞内生长所必须。
军团菌属中的菌株特异效应蛋白
在 608 个 LEOGs 中,258 个 LEOGs 为菌株特异的,即仅在一个军团菌种中存在。其中,188 个 LEOGs与军团菌基因组中的其它蛋白显示局部的序列相似性(E-value < 1×10-4),70 个LEOGs 与军团菌基因组中的其它蛋白无序列相似性。
效应蛋白基因的获得和丢失
结合进化树分析效应蛋白基因的获得与丢失,结果显示军团菌属中的效应蛋白库处于动态变化中,不同分支的效应蛋白获得与丢失的速率不同。进化关系上比较接近的种更倾向于具有相似的效应蛋白数据集,见图4。对这些经历大量的基因水平转移效应蛋白的进化分析结果表明,效应蛋白基因的基因水平转移事件更倾向于发生在进化关系接近的种间。
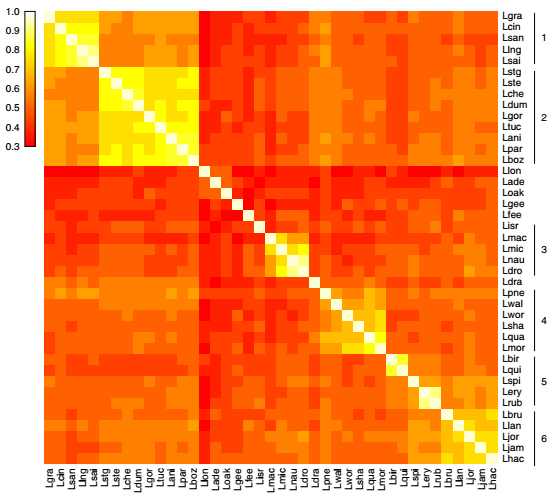
图4 假定效应蛋白池在军团菌属的比较
效应蛋白的共线性和协同进化
在一些案例中,在基因组中处在相近位置的效应蛋白编码基因在宿主细胞中往往协同起作用。研究者分析了多个基因组中连接在一起的效应蛋白编码基因(效应蛋白之间相邻不超过 5 个ORFs),总共找到 143 对效应蛋白编码基因至少在 2 个基因组中邻近,其中,51对效应蛋白编码基因组在至少5个基因组中邻近。进一步分析这些ORFs的共进化情况,发现这些效应蛋白的编码基因共同获取或者丢失的情况远远高于偶然几率,最终确定19对效应蛋白编码基因是共进化的,并且协同发挥作用。
效应蛋白进化的主因:结构域重组
之前的研究报道,L. pneumophila的效应蛋白包含大量来自真核生物的结构域和效应蛋白特异的结构域。采用两种方法对效应蛋白的保守结构域进行分析:与结构域数据库中的已知结构域进行比对分析;对效应蛋白直系同源物的保守区域分析。结果显示,在56%的假定蛋白中检测到了保守的结构域。99个结构域可通过上述两种方法同时检测到,其中 53 个结构域功能已知(大部分来自于真核生物),46 个结构域保守蛋白,功能未知。研究者发现不同的蛋白质结构经常分享相同的结构域。将蛋白质的结构通过共有的结构域进行连接形成网状结构进行可视化,节点为共有的结构域,发现锚蛋白重复结构域、磷脂酰肌醇-4-磷酸结合域(PI4P)和富含亮氨酸的重复单元(LRR)结构域为最大的蛋白结构连接区域,见图 5。通过PI4P结构域和LED006对编码效应蛋白的基因进行预测分析得到效应蛋白编码基因,具体结果见图6。上述结果验证了军团菌效应蛋白功能多样性的原因以及结构域重组对于军团菌共进化的重要作用
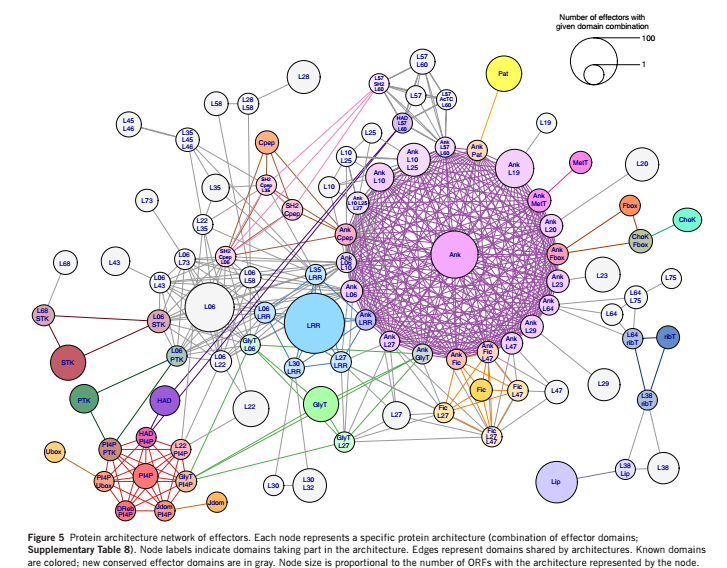
图 5 效应蛋白的结构网络
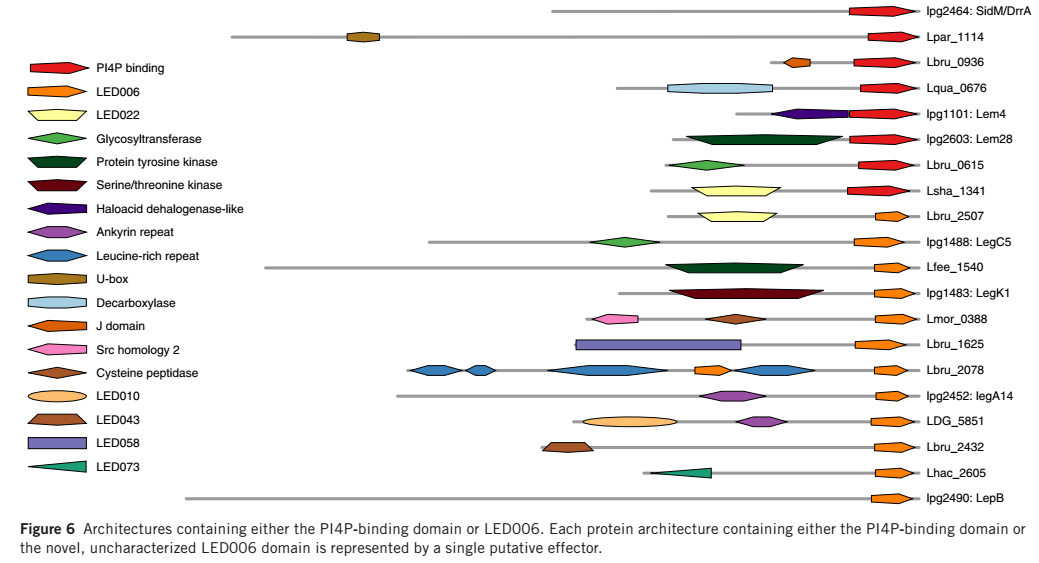
图 6 包含PI4P或LED006结构域的蛋白结构
结论:
在军团菌可以导致严重的疾病大背景下,通过高通量测序完成了38株军团菌的框架图,并针对其致病性进行深入分析。研究了与致病性相关的分泌系统和效应蛋白,发现编码分泌系统的基因较为保守,而编码效应蛋白的基因差异较大,只有7个效应蛋白编码基因为军团菌属核心共有基因。进一步分析效应蛋白多样性的原因发现结构域重组是其多样性的原因,而结构域在不同的军团菌中可以共有。
文章亮点:
选材好:选取的材料为人致病菌,受关注程度高;
立足点好:选取了 38 个军团菌属的菌株进行测序,故事内容主要是讲其致病基因,即效应蛋白和分泌系统;
会讲故事:研究者有浅入深,层层递进,完整地阐述了一个故事。


















