

2017-12-13
病原菌需要迅速调整其毒性和适应症,以防止被宿主消灭。到目前为止,关于潜在适应过程的驱动致病机制大多是都是体外研究实验,从而忽略了宿主诱导刺激对入侵病原体作用的真正复杂性。在本研究中,我们开发了一种实验方法,可以同时监测宿主和侵染的细胞外病原体的转录变化。利用该工具可以发现许多与炎症、穴位反应、凝血活动和过渡金属离子隔离有关的宿主转录本的改变,并强调免疫反应主要是浸润中性粒细胞和混合TH17 / TH1反应。因此,病原体的反应主要是为了抵抗吞噬细胞的攻击。肺结核菌上调了抗吞噬细胞III分泌系统(T3SS)的基因的表达,并诱导了对抗中性白细胞诱导离子剥夺、自由基应激和营养抑制的功能。对毒力影响最大的是碳储存调节剂(Csr)系统的丢失,对宿主细胞T3SS表达上调至关重要。综上所述,互作转录组测序为发现感染特异性刺激、诱导宿主和病原体反应以及潜在的调节过程提供了有力的工具。
组织互作转录组结果
如图1所示,(A)组织互作转录组结果显示,雌性BALB/c小鼠口服感染2×108 cfu肺结核菌IP32953(感染)或1×PBS(未感染)。体外RNA-seq实验所用IP32953为25°C或37°C指数期或者平台期的菌体。(B)基因组圈图可视化显示了IP32953染色体(NC_006155.1)和pYV质粒(NC_006153.2)体外和体内转录组数据。(C)从未受感染和感染的Peyer斑块的RNA池中,对3重复的小鼠RNA-seq数据进行了分析并制作热图。(D)主要成分(PC)分析均值为read数--来自结核菌IP32953核心基因组和pYV病毒质粒体外和体内转录组实验。
肺结核菌感染宿主反应
如图2所示, (A)DESeq2分析未受感染和受感染的Peyer的斑块RNA获得的火山图。(B)前50富集和前50减少的宿主热图。(C)免疫介导的外部组织因子依赖性凝血级联基因的活化。在IP32953的侵染过程中,受感染的Peyer斑块中,外来的凝血级联转录本已被高度浓缩。纤维蛋白凝块通常是由细菌抗原(如LPS)诱导的,以诱捕入侵细菌并阻止进一步的传播。此外,通过中性细胞特异性胶原酶MMP8刺激外源性凝血通路,激活TFPI的组织因子。
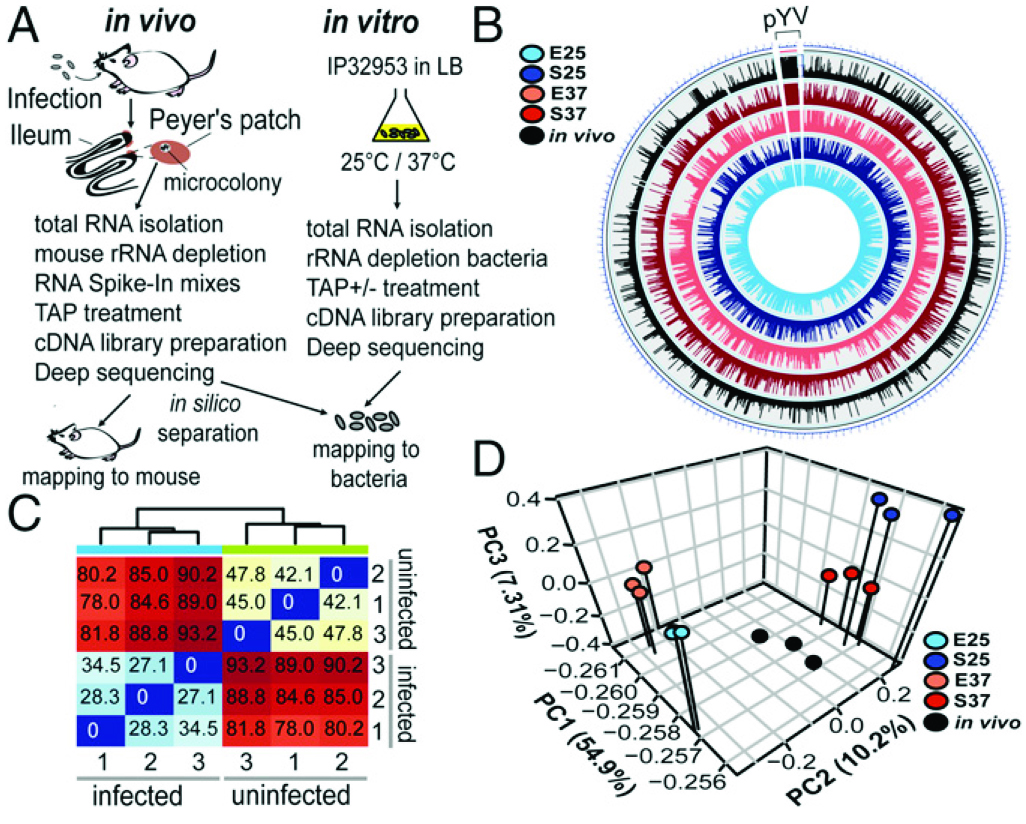
图1组织互作转录组流程和报告。
毒力因子表达及宿主细菌应激反应
如图3所示,细菌基因表达分析揭示了特定感染的代谢适应性。(A)维恩图展示了细菌侵染后的小鼠显著改变的转录本(log2fc≥1;log2fc≤−1;P≤0.05);与4种体外实验结果相比,体内实验有283个转录本发生了显著变化,包括在一个条件下富集的和在另一种情况下衰竭。在这283个转录本中,116个被持续富集,62个在体内持续衰竭。(B)在感染期间严格富集 (红色)或衰竭(蓝色)的选定细菌的基因热图。(C)结核杆菌中心碳代谢。在体外和体内培养的细菌中,转录模式有了显著变化。在感染过程中,富集的转录本用红色表示,而减少的转录本用蓝色表示。
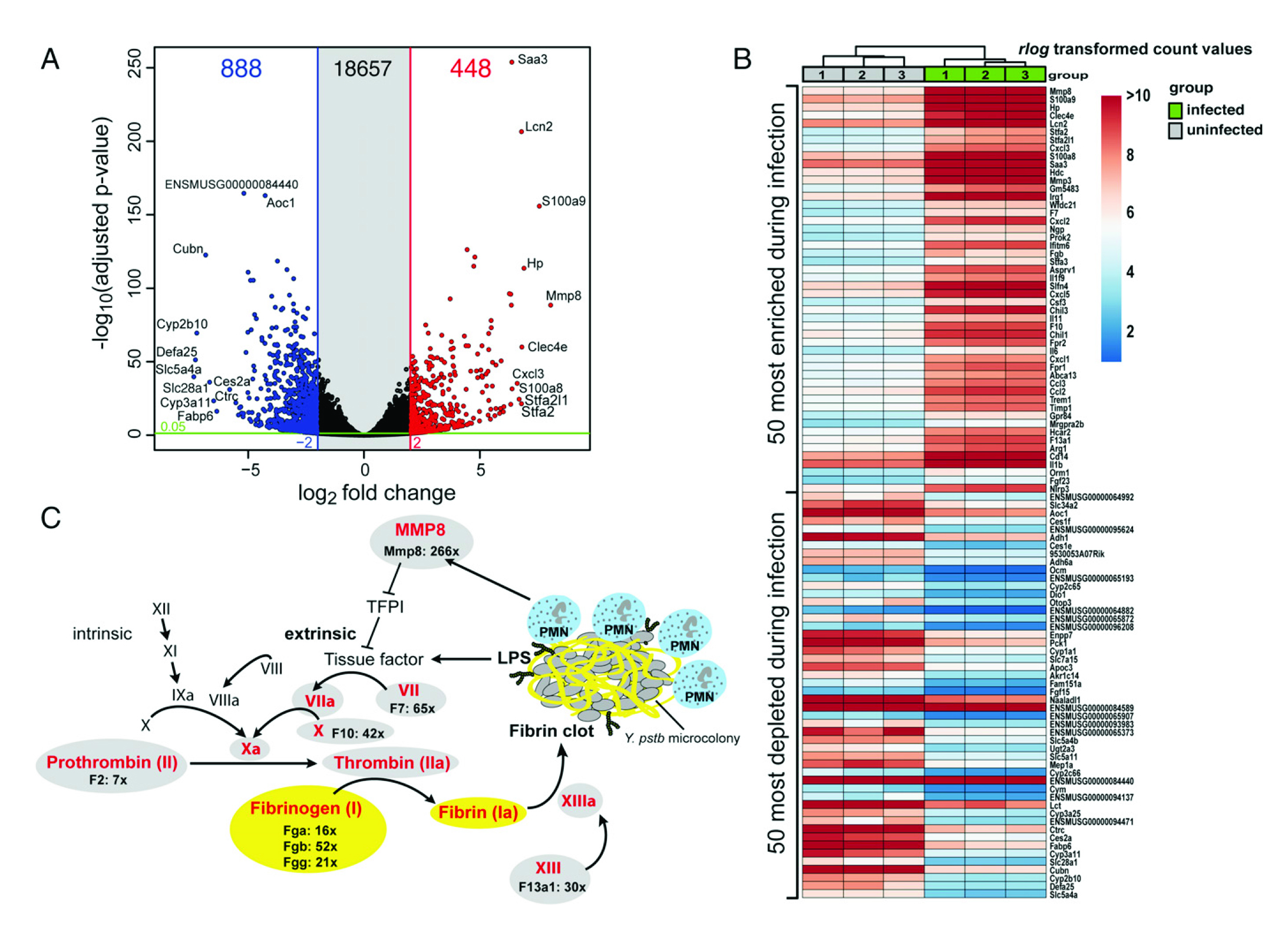
图2宿主对肺结核菌的转录反应
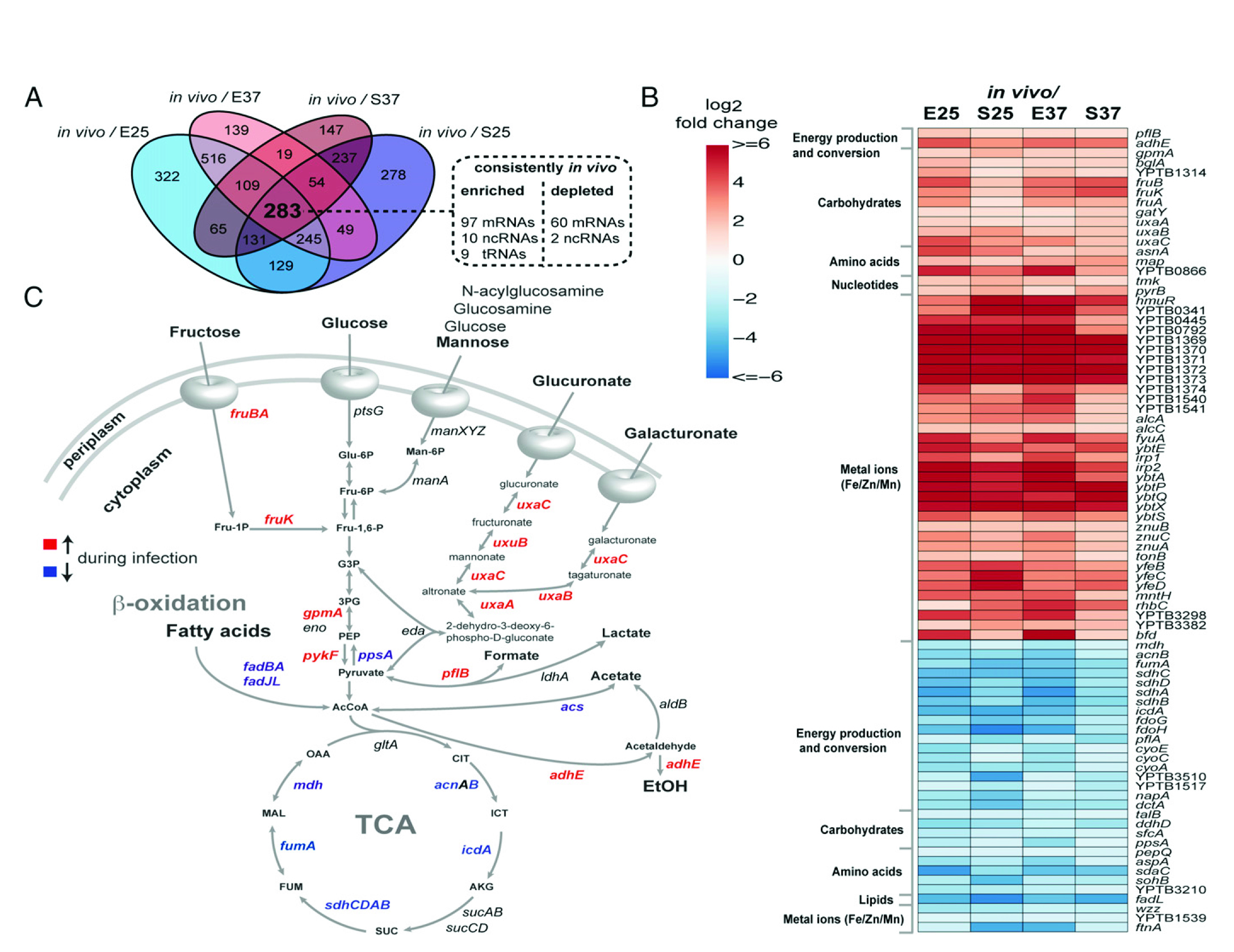
如图3细菌基因表达分析
淋巴组织的生长改变了保守的代谢ncRNAs的数量
图4显示,高度保守的细菌ncRNAs有助于发病。(A)反式编码的细菌ncRNAs的富集的热图(红色)和衰竭的热图(蓝色)。 (B) sgrS、ryhB1、ryhB2和glmY loci的rna - seq分析的read覆盖率说明。数据按照惟一比对的reads进行了规范化。(C)结核菌野生型IP32953和同基因的ΔsgrS在LB培养基培养37°C,添加和未添加αMG 1%,磷酸葡萄糖胁迫。(D)两个独立的BALB/c小鼠组(每组2×n = 5)口服等量混合感染107 cfu IP32953(WT)和同基因的突变YPIP56(ΔsgrS/ryhB1/ryhB2/glmY)。在Peyer斑块(PPs)中,肠系膜淋巴结(MLNs)和3天后器官的组织样本被添加不同的接头。

图4 高度保守的细菌ncRNAs
图5所示,CsrA对T3SS的毒力和介导其表达至关重要。(A)小鼠口服感染2×108 cfu结核菌野生型YPIII或同基因的ΔcsrA菌株。在两个独立的实验中每组每个基因型老鼠2×10只,实验12 d监测老鼠的存活率。(B)BALB/c小鼠口服感染107 cfu结核菌YPIII WT或同基因的ΔcsrA菌株。小鼠感染3 d后被杀,确定均质Peyer斑块(PPs)、肠系膜淋巴结(MLNs)、肝脏和脾脏内的细菌数目。数据来自两个独立实验(2×5老鼠/组)。(C)TCA-沉降上层清液或(D)Westernblots:全细胞提取物的结核菌YPIII WT和同基因的ΔcsrA菌株和没有补充csrA +质粒pAKH56生长在37°C缺少Ca2 +的培养基。(C)考马斯亮蓝染色或(D)Western blotting鉴定。核酸相关蛋白H-NS被用作阴性对照。
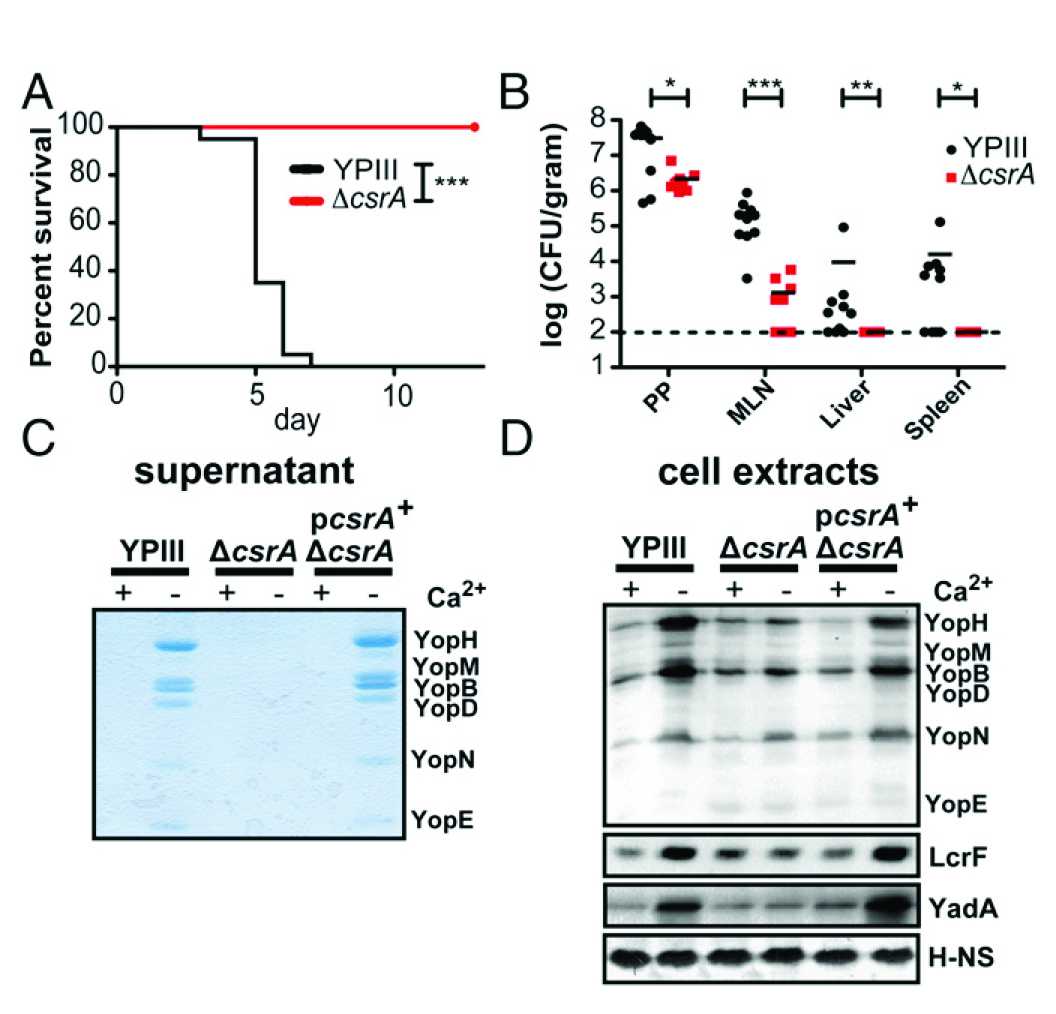
图5 表型实验
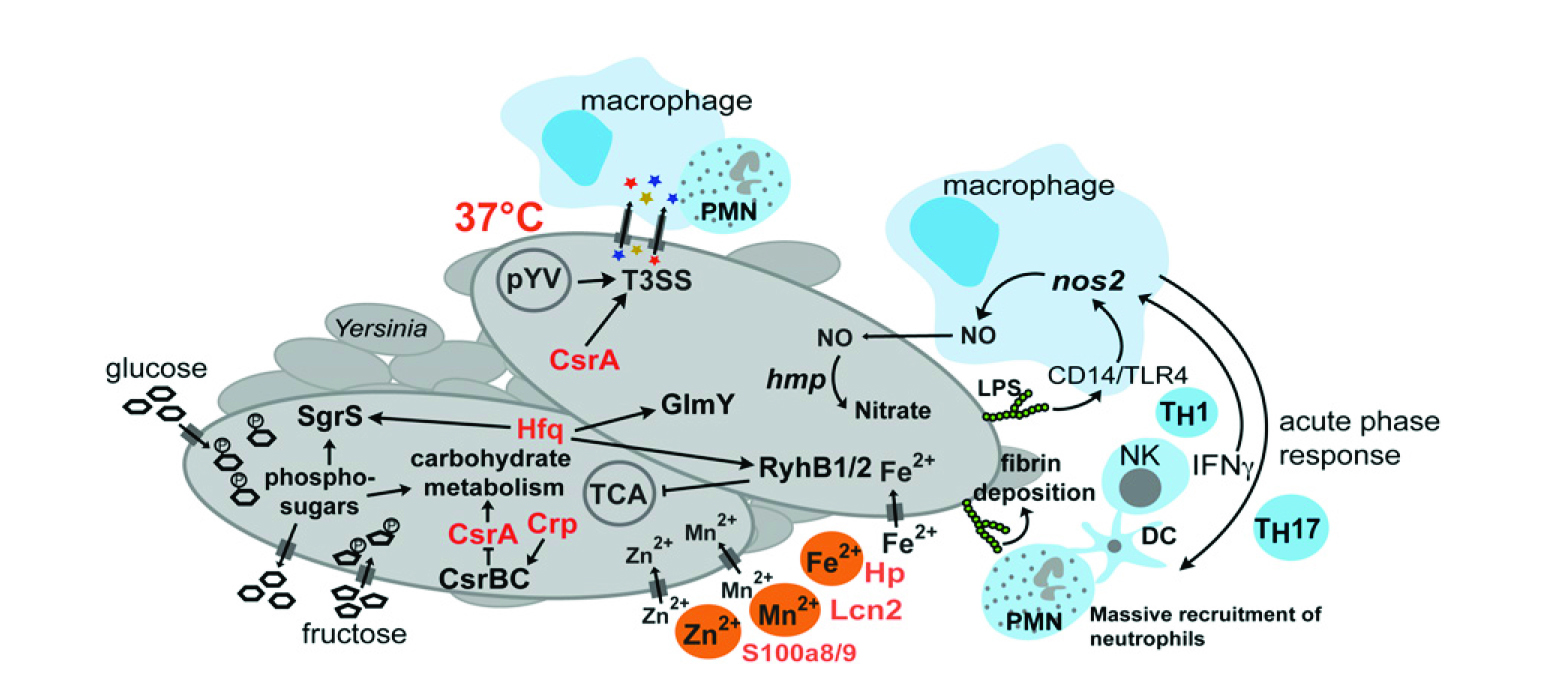
图6 结核菌-宿主转录组变化
如图6所示,结核菌-宿主转录组的特定感染变化。在Peyer斑块的侵染过程中,对结核菌和宿主的复杂转录本适应性的示意图。最受影响的炎症和急性期反应,代谢和毒力的适应,并识别不同的细菌核素从而影响发病机制。
综上所述,以上结果证明了组织互作转录组方法的有用性,可以得出与病原体生物学和宿主反应相关的有意义的结论。通过免疫细胞迁移和细菌致病机理的数据整合获得的RNA剖面信息可以深入研究复杂的宿主-病原体相互作用网络和发病机理。它进一步创造了识别控制病原体侵染的关键过程的可能性,并增加了识别抗生素治疗目标的潜力。


















