

2017-07-19
SlARF10,番茄生长素响应因子,受Sl-miR160的转录调控。通过表达SlARF10(mSlARF10),发现其小叶叶片比较窄,叶片上有较大的气孔但密度比较低。35S:mSlARF10-6植株和野生型WT相比, 其窄型离体叶片失水较多。研究发现了之前未识别到的一个叶片失水调控因素,以及在这个重要过程中生长素和ABA信号协调作用的互作网络关系。miR160调控ARF10,来维持叶片中的水平衡,以保证正常的植株生长和环境适应性。
1、研究发现在35S:mSlARF10-6植株和野生型WT中,实际的失水量和计算的气孔的失水量是不一致的,表明他们存在导水率差异。过表达mSlARF10可能会通过蒸腾作用增加了转基因植株的叶片的失水量。35S:mSlARF10-6叶片的导水率要比WT中显著提高(图1b)。结果显示mSlARF10过表达不仅改变叶片性状,而且影响导水率。
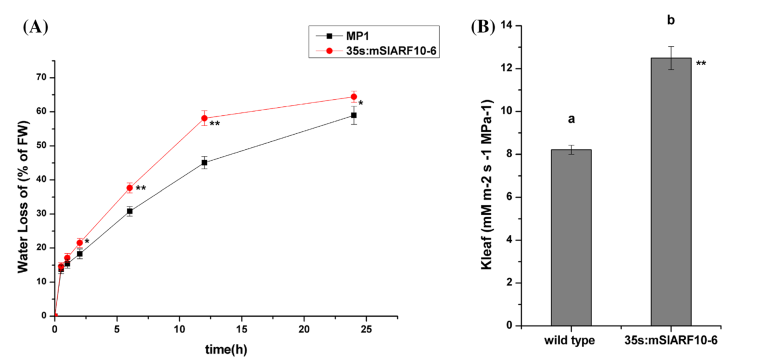
图1,过表达mSlARF10增加了叶片失水。a:叶片失水率,b:叶片导水率
2、用ABA(脱落酸)和HgCl2做预处理,验证在35S:mSlARF10中有较高的导水率,这和其有较大的气孔和较高活性水通道蛋白相关(AQPs)。ABA处理的条件下,35S:mSlARF10-6有着较高的敏感性,气孔会迅速关闭。在WT的0-24h的脱水处理中,内源性ABA的积累逐渐增加。ABA敏感性实验检测SlARF10是否和ABA信号传导相关。在35S:mSlARF10-6和WT叶片中,ABA处理可能减少了失水(图2)。ABA处理的35S:mSlARF10-6中叶片的失水很大程度的下降,将近20%。ABA处理的WT的叶片失水量在脱水处理6h后,比脱水处理6h前还低。35S:mSlARF10-6中失水速度比WT中要高,但是ABA处理后,在脱水处理6h前要比WT中要慢很多。这表明在早期的脱水处理中,ABA在35S:mSlARF10-6叶片中对失水的抑制作用要比WT中要大。
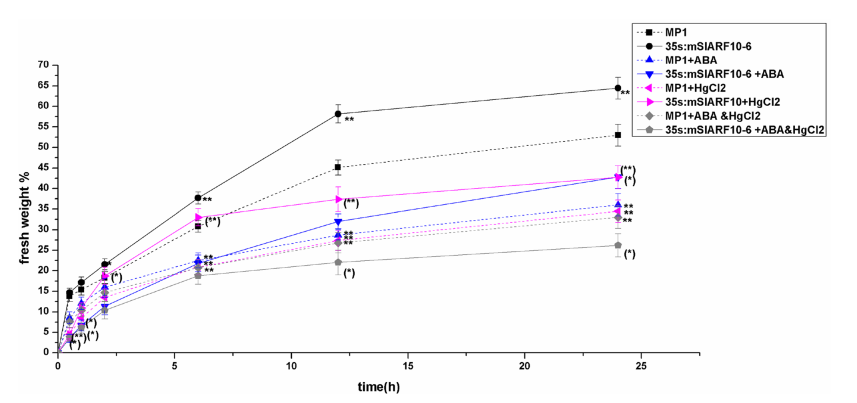
图2,不同处理条件下,叶片的失水率
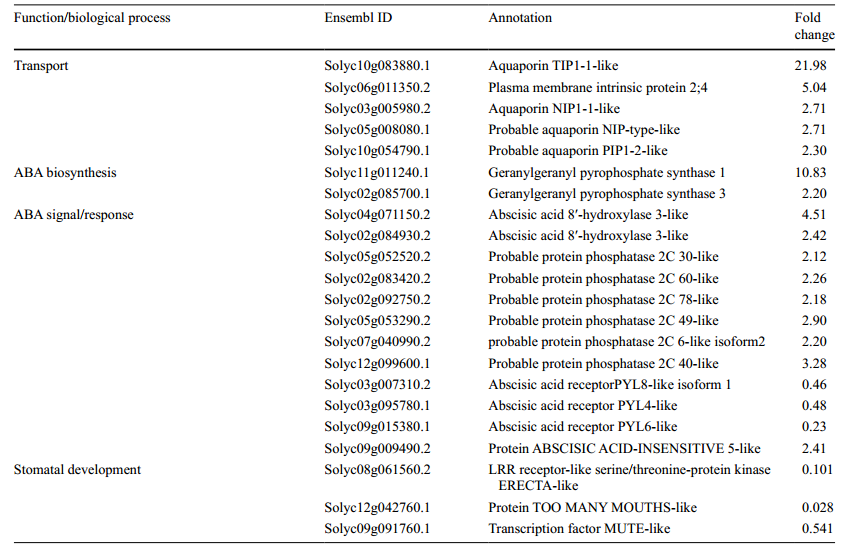
3、RNA测序表明5种AQP相关的基因,14种ABA合成/信号传导基因和2种气孔发育基因,在35S:mSlARF10-6植株中明显被改变,这一结果被qRT-PCR验证。用WT和35S:mSlARF10-6的15个叶片做RNA测序,检测35S:mSlARF10-6叶片对整体ABA合成,信号传导,和AQP基因表达的影响。结果发现在35S:mSlARF10-6样本中发现5个显著上调的QP基因,2个ABA合成和12个信号/响应显著上调或下调的基因(表1)。
表1,叶片中差异性上调的AQPs,ABA和气孔发育相关的基因
4、实验发现,SlARF10通过调节气孔开合度来缓和失水现象,增强了ABA合成/信号响应;也通过影响气孔的形成和AQP的表达来影响水的运输,二者一起协调控制番茄叶片的失水。
构建互作反应的网络图,说明miR160在ARF10在植物水压环境中的调控作用中的精确作用(图3)。该模式说明ABA-植物生长素相互作用对控制失水的影响。当生长素信号被激活,Aux/IAA被分解,ARF10被高水平转录。尽管ARF10可以通过ABA合成基因活跃的表达增强ABA含量,通过ABI5表达增强ABA信号,最后导致干旱是气孔关闭。适当表达水平受miRNA160调控的ARF10,可能在叶片发育中维持叶片水平衡和适应水压中有着重要的作用。
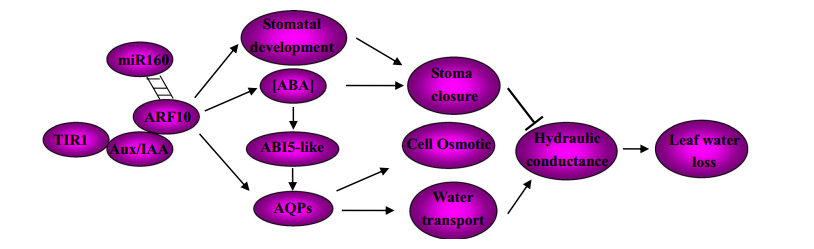
图3,针对miR160调控的ARF10在对叶片失水的响应中起的作用提出了一种互作模式
结论:
1、 mSlARF10的过表达导致叶片变窄和更高的失水率
2、 mSlARF10的过表达通过影响ABA合成和信号传导调控气孔关闭
3、 mSlARF10的过表达诱导叶片表皮的AQP表达
4、 受miR160调控的SlARF10控制叶片中生长素和ABA对失水现象的响应


















