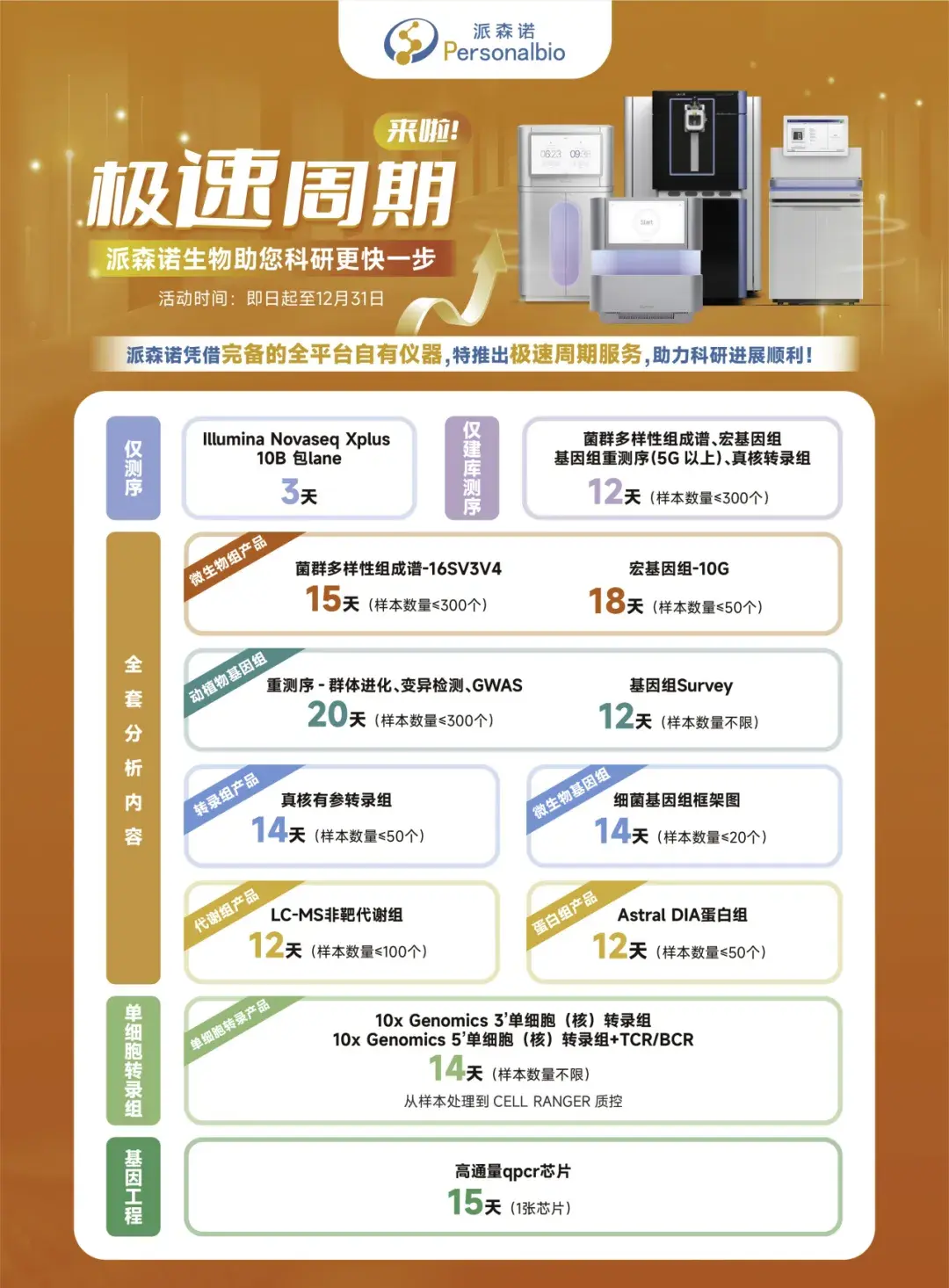
2024-11-08

期刊:Developmental Cell
影响因子:10.7
研究背景
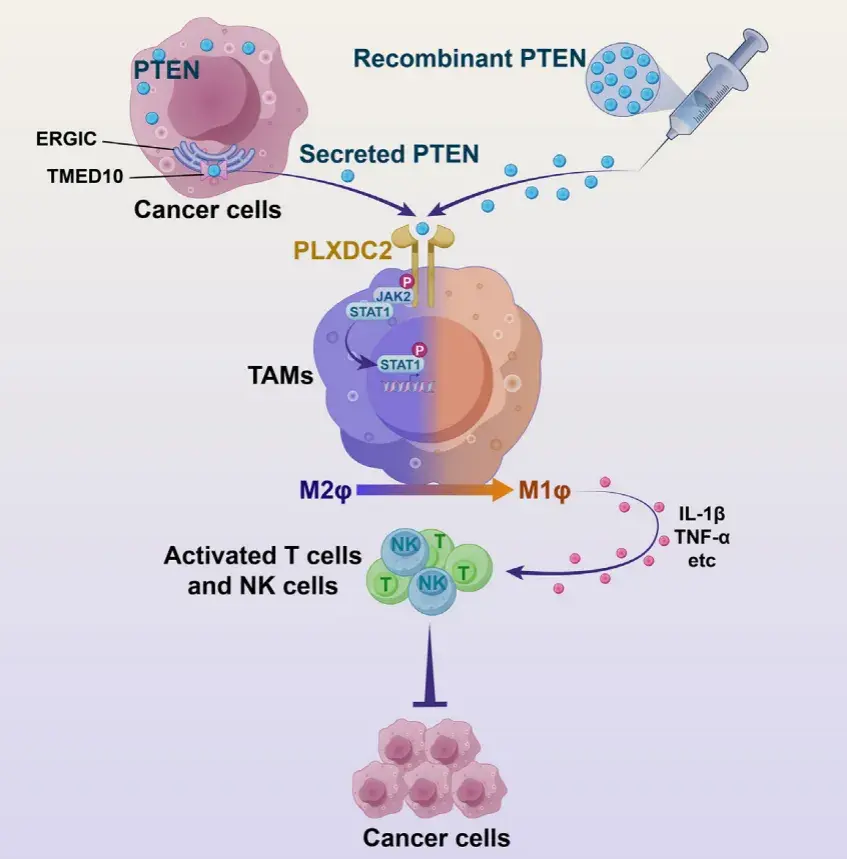
PTEN 是人类癌症中最常见的肿瘤抑制因子之一,其功能的缺失与多种癌症的发生、发展以及对免疫治疗的耐药性密切相关。然而,已有研究主要集中在 PTEN 的细胞内功能,对其在肿瘤微环境(TME)中的免疫调节作用了解有限。特别是,PTEN 缺失的肿瘤通常表现出免疫抑制微环境,增加了肿瘤对免疫疗法的抵抗力。近年来,研究表明 PTEN 可以通过一种非传统的蛋白质分泌途径分泌到细胞外,然而其在肿瘤免疫中的具体机制尚未明确。2024年8月27日,上海交通大学医学院附属仁济医院/海南医科大学陈国强院士团队在Developmental Cell上发表了文章Secreted PTEN binds PLXDC2 on macrophages to drive antitumor immunity and tumor suppression,揭示了PTEN可通过非经典蛋白分泌通路THU以可溶形式分泌到细胞外,并通过重编程肿瘤免疫微环境,实现抗肿瘤免疫的激活与肿瘤抑制。派森诺生物为该研究提供了单细胞转录组测序服务。
结 果
01 、PTEN 通过 TMED10 通道蛋白分泌途径分泌

作者在多种细胞系的培养上清液中都发现了PTEN蛋白及其他已知的分泌蛋白,如HMGB1 和Gal-1。而293T 细胞上清液中缺少GM130,这排除了上清液中的PTEN蛋白来自死细胞的可能性。另一方面,从PTEN 敲除的小鼠胚胎成纤维细胞和PTEN缺陷的U251胶质母细胞瘤细胞上清液中未检测到PTEN,进一步证明 PTEN来自活细胞。ELISA试验表明,293T细胞在1小时内分泌了450 pg PTEN ,而通过CRISPR-Cas9 系统敲除 PTEN 则消除了该分泌。这些结果表明,细胞内的 PTEN 蛋白可被分泌到细胞外。
考虑到有报道称 PTEN 是通过外泌体分泌的,作者还探讨了外泌体介导 PTEN 分泌的可能性。为此,作者收集了与293T细胞孵育1小时和24小时的培养基,通过超滤分离和TEM分析,发现外泌体中PTEN的含量有限,证明外泌体不是 PTEN 分泌的主要途径。
然后作者进一步排除了 PTEN 通过常规蛋白质分泌途径分泌的可能性,因为布雷菲德菌素 A 可阻断分泌蛋白从内质网到高尔基体的运输,但对293T 细胞分泌内源性 PTEN 没有影响。这与使用 SignalP 预测的 PTEN 蛋白缺乏典型分泌信号的事实一致。为了找到 PTEN 分泌途径的线索,作者转向 PTEN 相互作用蛋白,发现PTEN与HSP90B1、TMED10和HSP90AB1相互作用,这些是THU通路的组成部分。敲除 TMED10发现可显著降低分泌的 PTEN 量,而过表达 TMED10 可进一步增加分泌的 PTEN 量。此外,HSP90AB1 或 HSP90B1 的耗竭会抑制 PTEN 的分泌,而它们的共同耗竭则显示出最强的效果。这些结果表明 THU 可能是 PTEN 分泌的主要途径。
为了巩固 PTEN 通过 THU 分泌的观点,作者发现PTEN与TMED10共定位在ERGIC上,并与TMED10中的类似区域结合,如同IL-1b。通过MEME-Suite网站分析,作者发现PTEN和IL-1b有共同基序,其中基序3对PTEN分泌至关重要。W274是指导THU分泌PTEN的关键氨基酸残基,W274突变几乎消除了PTEN与TMED10的相互作用。PTEN的W274在物种中高度保守。综上,TMED10通过识别PTEN的W274来指导其通过THU通路。
02、细胞外可溶性 PTEN 抑制肿瘤生长和免疫反应
2.1 细胞外可溶性 PTEN 抑制肿瘤生长
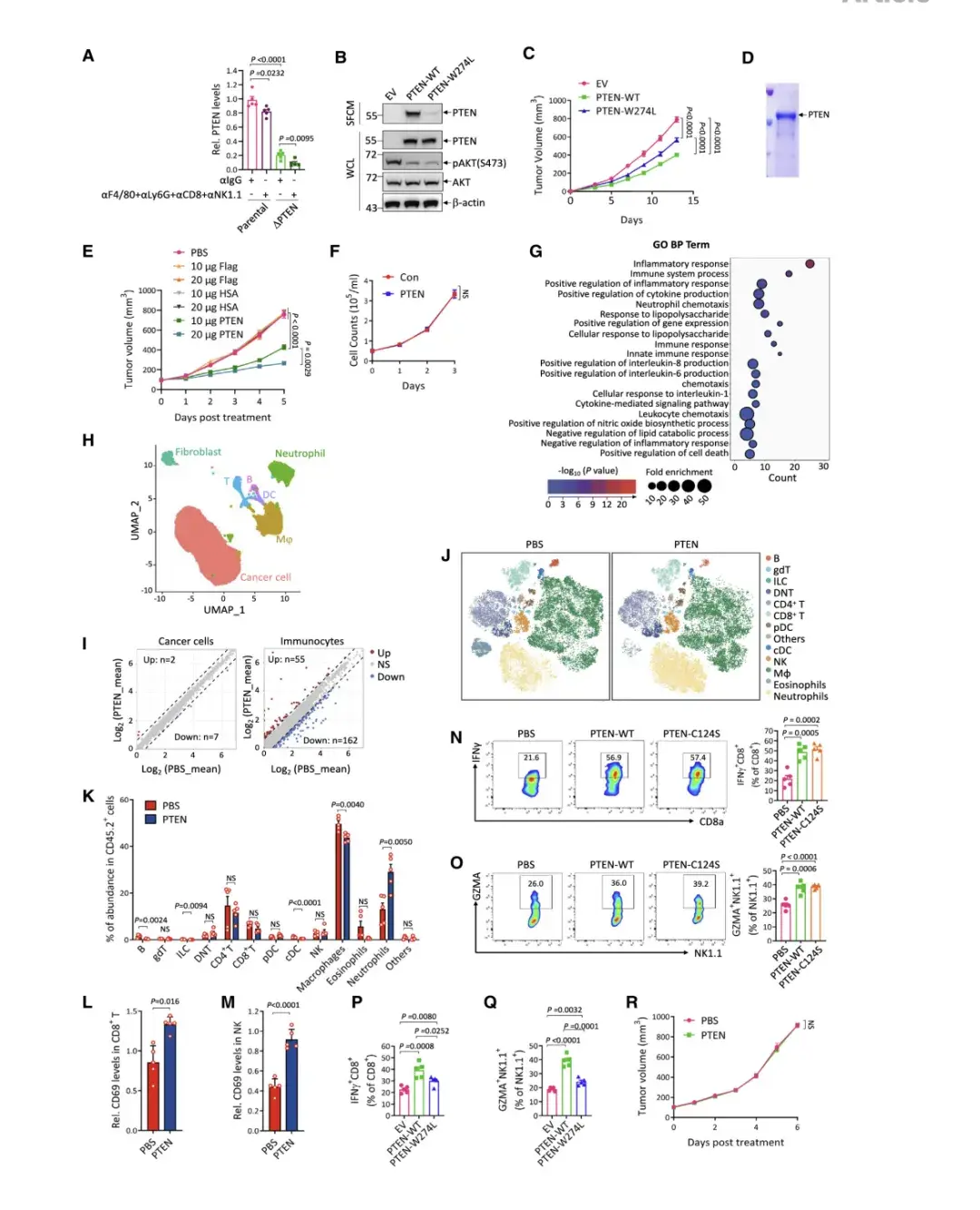
作者用ELISA试验检测B16-F10小鼠黑色素瘤细胞肿瘤间质液(TIF)中的PTEN蛋白,发现PTEN以22 ng / mL的浓度存在于TIF中,这验证了TME中细胞外PTEN的存在。而后为了确定TIF中PTEN的来源,比较亲本B16-F10细胞和PTEN敲除的B16-F10细胞系产生的TIF,发现后者中PTEN减少了约80%,说明肿瘤细胞是TME中分泌PTEN的主要来源。
为了评估分泌型 PTEN 在 TME 中的作用,将 PTEN 敲除的 MC38 细胞 (MC38-DPTEN)进行野生型 PTEN (PTEN-WT) 或含有 W274L 突变的 PTEN 突变体 (PTEN-W274L)再表达,发现PTEN-W274L 的分泌效率低于 PTEN-WT,但它们对 AKT 磷酸化 (Ser473) 的抑制强度相似 ,这表明 W274L 突变选择性地影响 PTEN 的分泌,但不影响其磷酸酶活性。然后,将这些细胞系在 C57BL/6J 小鼠中进行皮下生长,结果显示与 PTEN-WT 相比,PTEN W274L 的肿瘤抑制作用受损 ,这表明分泌型 PTEN 可能在肿瘤抑制中发挥作用
为了进一步评估分泌型 PTEN 在肿瘤抑制中的作用,作者尝试直接用 PTEN 蛋白治疗肿瘤。
与所有对照组相比,PTEN 蛋白治疗表现出显著的剂量依赖性肿瘤抑制作用。细胞外 PTEN 的肿瘤抑制作用也可以在 MC38-DPTEN 细胞产生的肿瘤中重现。值得注意的是,纯化的 PTEN 蛋白携带一种使其磷酸酶活性失活的突变(PTEN-C124S),其对肿瘤生长的抑制程度与 PTEN-WT 相似。此外,腹膜内注射纯化的 PTEN 也显著抑制了B16-F10-DPTEN 肿瘤的生长。总之,细胞外可溶性PTEN以磷酸酶活性非依赖的方式发挥肿瘤抑制作用。
2.2 细胞外 PTEN 引发抗肿瘤免疫反应
与体内抑瘤作用相反,细胞外 PTEN 治疗不影响 B16-F10DPTEN 细胞的体外生长,这暗示它可能作用于TME。事实上,当通过 RNA 测序分析 PTEN 治疗的 B16-F10-DPTEN 肿瘤时,对 104 个失调基因的 GO 分析显示,它们主要富集在参与免疫反应的途径中。特别是,干扰素 a 和 IFNg 反应的强烈激活表明 PTEN 治疗会在肿瘤中引发抗肿瘤免疫反应。此外,作者对 PTEN 治疗和未治疗的 B16-F10DPTEN 肿瘤进行单细胞 RNA 转录组测序 (scRNA-seq)。结果显示,与转录组几乎未受干扰的癌细胞相比,免疫细胞的转录组发生了显著变化,表明 PTEN 治疗优先改变了 TIME。
接着,作者使用质谱流式细胞术对接受 PTEN 治疗和未接受 PTEN 治疗的 B16-F10-DPTEN 肿瘤中的免疫细胞进行了分析。中性粒细胞总数变化最为显著,巨噬细胞总数略有改变,而 CD4+ /CD8+ T 细胞和 NK 细胞的总数未受到影响。然而,在接受 PTEN 治疗的肿瘤中,表达活化标志物 CD69 的 CD8+ T 细胞和 NK 细胞的比例显著增加。通过对 B16-F10DPTEN 肿瘤以及 MC38-DPTEN 肿瘤中的 IFNg 阳性 CD8+ T 细胞和颗粒酶 A (GZMA) 阳性 NK 细胞进行流式细胞术分析,进一步证实了 PTEN-WT 或 PTEN-C124S 治疗增强了 CD8+ T 细胞和NK细胞的活化。与此同时,MC38-DPTEN 肿瘤中 PTEN 分泌的丧失导致了免疫抑制表型。结果表明,细胞外 PTEN 可增强抗肿瘤免疫力。此外,当用 PTEN 治疗在免疫缺陷 NCG 小鼠中生长的 B16-F10-DPTEN 肿瘤时,未观察到对肿瘤生长的明显影响,这表明细胞外 PTEN 以免疫反应依赖的方式抑制肿瘤生长
由于之前已证实 PTEN 无法在没有递送剂的帮助下进入细胞,作者推测它是否与 TME 中的免疫细胞表面结合。作者从 PTEN 治疗的 B16F10-DPTEN 肿瘤中分离出上述 PTEN 调节的免疫细胞(巨噬细胞、中性粒细胞、T 细胞和 NK 细胞),并测试了 PTEN 在其表面的存在。在四种类型的免疫细胞中,只有巨噬细胞被 PTEN 标记。通过将 PTEN 蛋白与从 C57BL/6J 小鼠脾脏中分离的免疫细胞进行体外孵育,可以确认 PTEN 在巨噬细胞结合中的偏好。此外,PTEN 以剂量依赖性方式与从骨髓来源的巨噬细胞(BMDM)和腹腔中分离的巨噬细胞结合。将 FLAG 标记的 PTEN 与未通透的 BMDM 一起孵育后,通过对 PTEN 进行免疫荧光染色,也可以观察到 PTEN 在 BMDM 表面上的存在,而未观察到细胞内信号。所有这些结果都表明 PTEN 与巨噬细胞表面结合。
03、细胞外 PTEN 使巨噬细胞极化为 M1 样表型
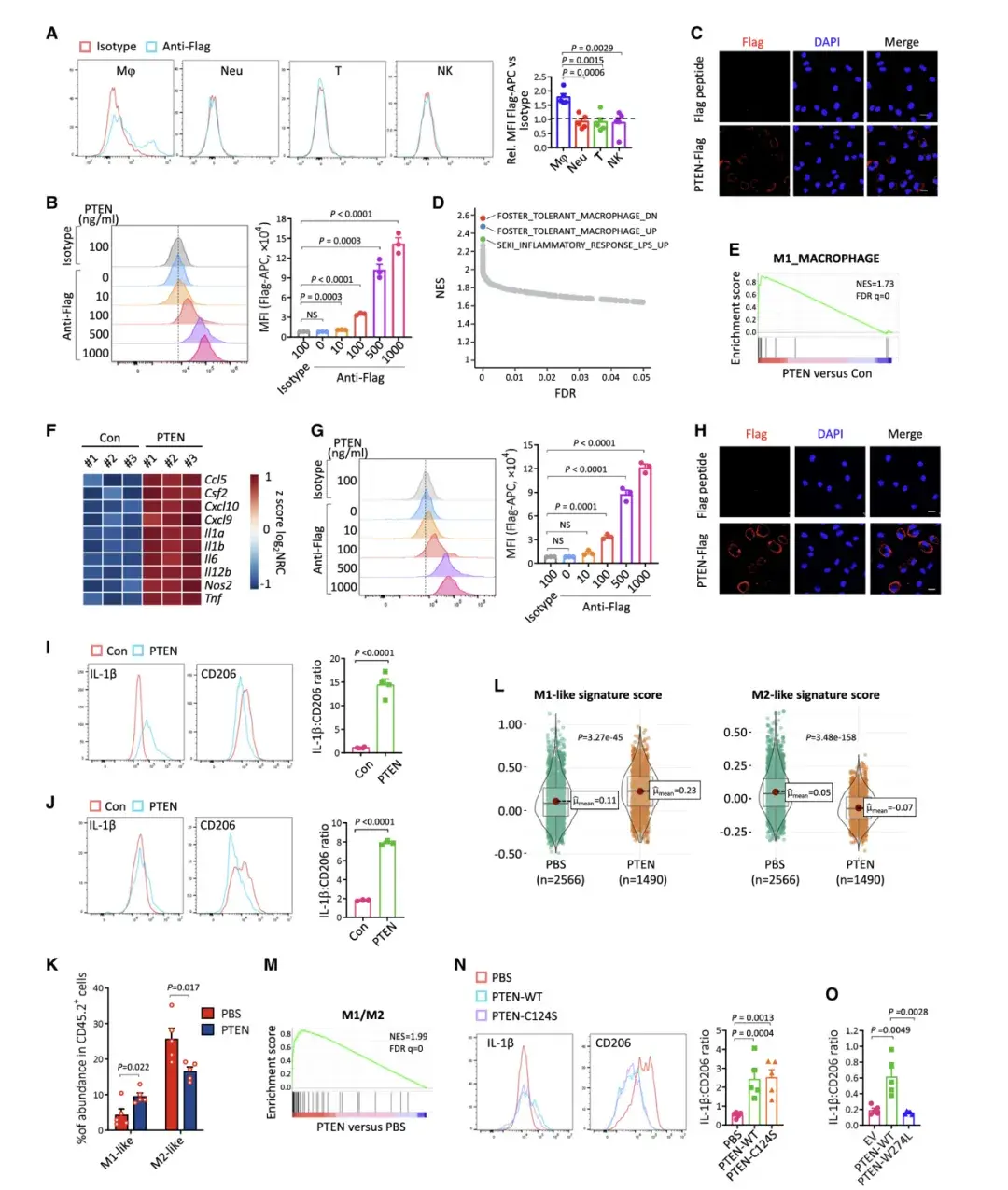
虽然 PTEN 治疗广泛改变了 BMDM 的转录组,但基因集富集分析 (GSEA) 表明,前三个上调通路都与脂多糖 (LPS) 刺激的信号传导有关。
LPS 是一种常规刺激物,可诱导 BMDM 的促炎(M1 样)极化, 这促使作者推测PTEN可能诱导BMDM发生M1样极化。而GSEA和M1标志物表达的增加支持了这一观点。这些结果表明,细胞外 PTEN 治疗使BMDM 极化为 M1 样表型。
TME中的巨噬细胞被称为肿瘤相关巨噬细胞 (TAM),它们与免疫抑制性 M2 极化巨噬细胞非常相似。为了研究PTEN对TAM的影响,研究者先用IL-4和IL-13处理BMDM诱导M2样极化。PTEN治疗增加了M1标志物的表达,降低了M2标志物的表达,并显著提高了IL-1b与CD206的表达比。这些结果表明,PTEN能将M2样巨噬细胞转化为M1样表型。
先前CyTOF数据显示 PTEN 治疗的肿瘤具有明显较少的 M2 样巨噬细胞,却有更多的 M1 样巨噬细胞。另外,作者利用scRNA-seq 数据在TAM中计算了单细胞水平上基于 M1 和 M2 基因特征的分数,PTEN治疗的肿瘤中分离的巨噬细胞显示出明显更高的 M1 基因特征分数,且M2基因特征分数低得多。RNA测序数据的GSEA分析也显示PTEN处理的肿瘤发生了从M2到M1的转化。流式细胞术分析表明PTEN治疗增加了巨噬细胞中IL-1b与CD206的表达比率,且这种作用不依赖于PTEN的磷酸酶活性。PTEN-WT在肿瘤中的重新表达增加了IL-1b与CD206的表达比率,而PTEN-W274L的重新表达则显示出大部分受损的作用。这些结果表明PTEN治疗使TAM极化偏向M1样表型。
04、PTEN 依赖巨噬细胞增强抗肿瘤免疫,抑制肿瘤生长
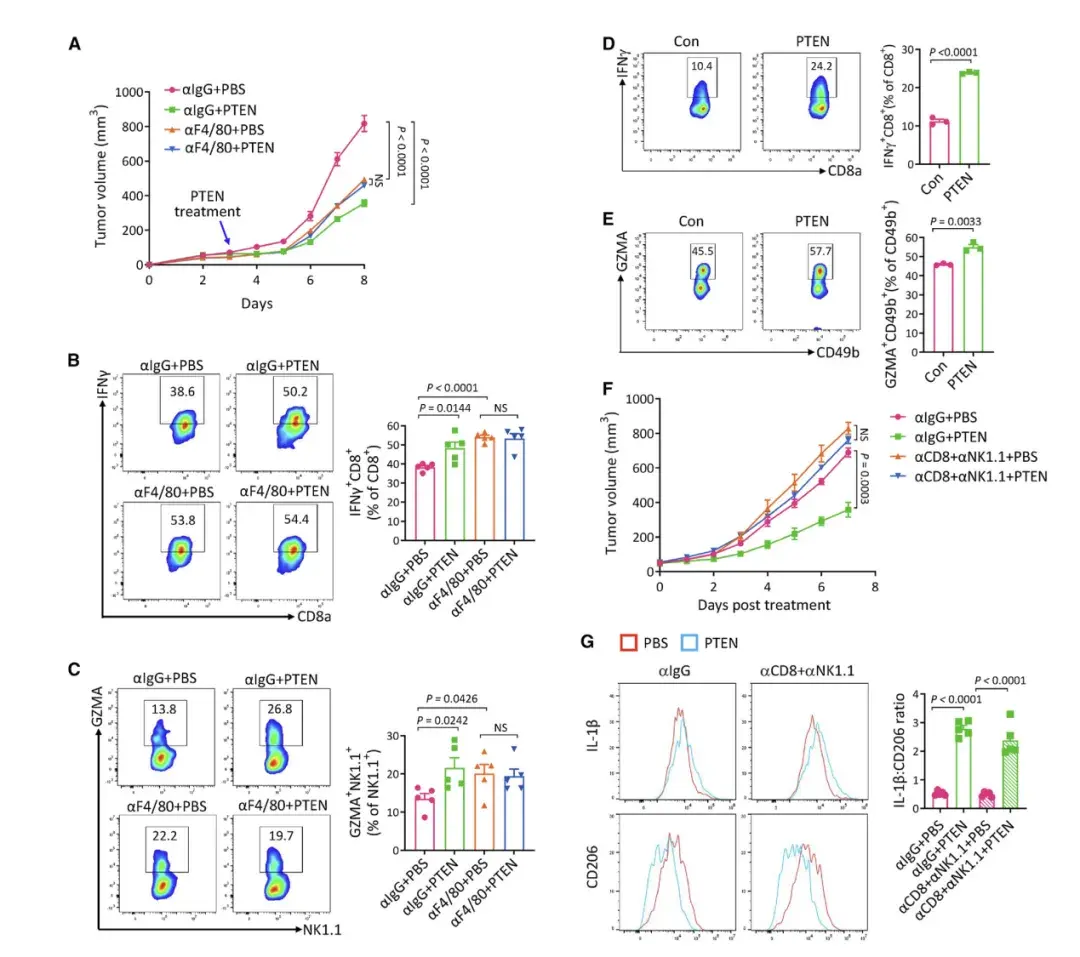
作者发现在缺乏PTEN的肿瘤中不再观察到PTEN治疗的抑癌作用,同时也不再促进CD8+ T和NK细胞的活化。此外,巨噬细胞的耗尽消除了PTEN 治疗在促进 CD8+T和NK细胞活化方面的作用,这表明 TAM 是介导PTEN治疗激活 CD8+T和NK细胞所必需的。这一观点在体外得到了进一步证实。所有这些结果表明巨噬细胞极化是PTEN治疗下CD8+T和 NK细胞激活的上游事件,它们的顺序发生介导细胞外PTEN的肿瘤抑制作用。作者通过CellChat软件分析scRNA-seq数据,发现SPP1-CD44轴在TAM抑制T细胞中起关键作用,并且PTEN处理显著影响TAM和T细胞之间的相互作用。尽管PTEN治疗增加了中性粒细胞的数量,但它们对于PTEN介导的肿瘤抑制并非必需。中性粒细胞的增加与巨噬细胞极化无直接关联。因此,PTEN治疗后中性粒细胞增加的机制和作用需要进一步研究。
05、PTEN 与巨噬细胞上的 PLXDC2 结合发挥抗肿瘤作用
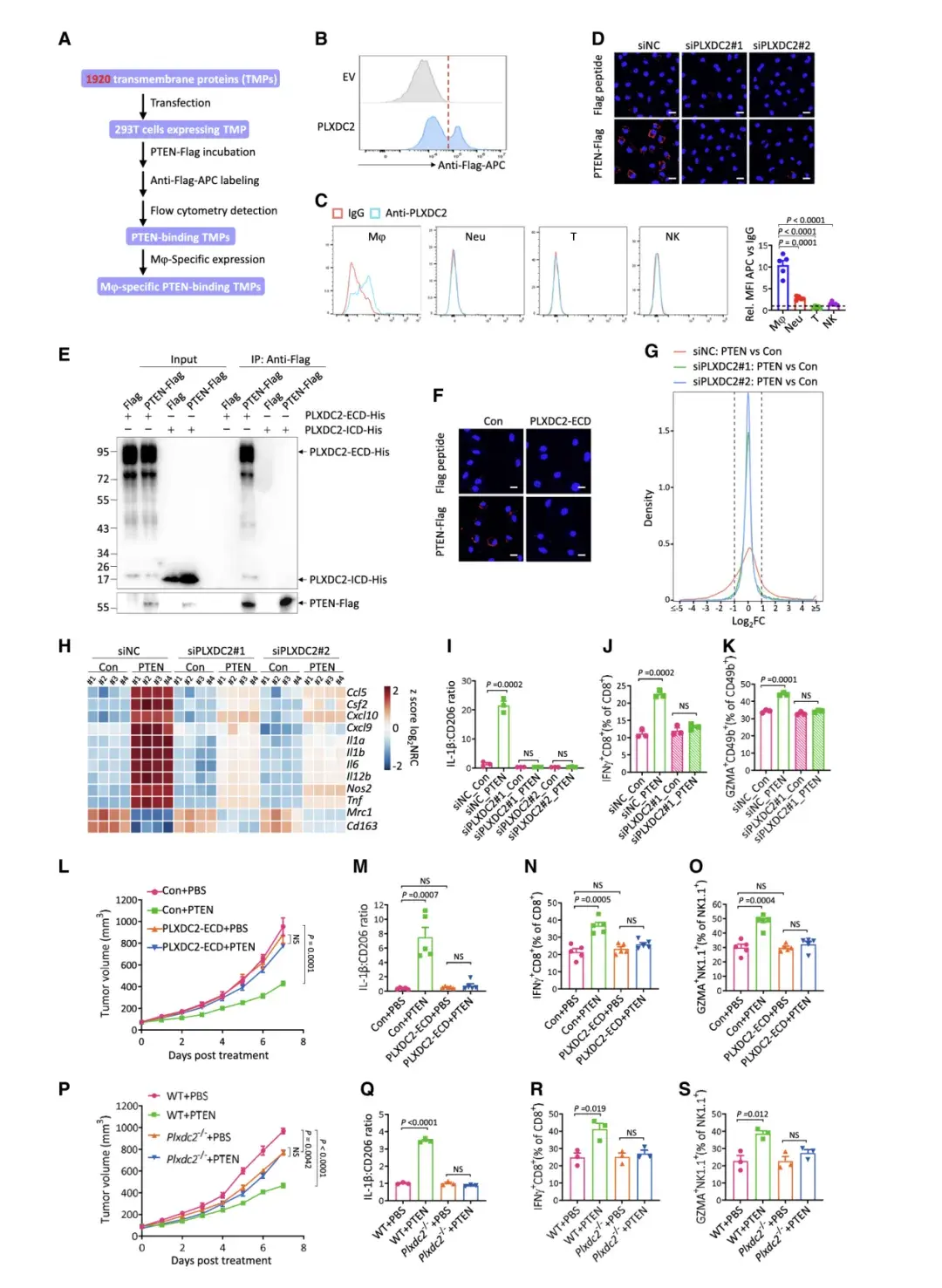
由于PTEN未进入巨噬细胞表面,而是优先与其结合,因此作者推测巨噬细胞存在特异性PTEN受体。为了识别该受体,作者通过数据库筛选出了PLXDC2。PLXDC2 特异性地表达在巨噬细胞的表面,但不表达在中性粒细胞、T细胞或NK细胞的表面,敲低PLXDC2在很大程度上阻止了 PTEN 与M2极化的BMDM结合。
最后,作者发现PTEN在体外直接与PLXDC2的细胞外结构域 (ECD) 相互作用,但不与细胞内结构域 (ICD) 相互作用,并且不与细胞中缺乏ECD的PLXDC2相互作用。将过度纯化的PLXDC2-ECD 添加到培养基中会消除PTEN与M2极化的BMDM表面的结合。这些结果都表明 PLXDC2 是一种介导 PTEN 与巨噬细胞表面结合的 PTEN 受体。
体外实验显示PTEN直接与PLXDC2的细胞外结构域相互作用。PLXDC2敲低显著改变了PTEN处理下M2极化BMDM的转录组,逆转了PTEN处理导致的基因变化。PLXDC2缺陷的BMDMs无法介导体外PTEN激活CD8+T和NK细胞的作用。使用PLXDC2敲低的巨噬细胞与B16-F10DPTEN细胞共植入小鼠,发现PTEN的肿瘤抑制作用在PLXDC2敲低组中显著减弱。骨髓特异性PLXDC2耗尽小鼠实验也显示,PTEN处理对这些小鼠产生的肿瘤无抑制效果。这些结果表明,细胞外 PTEN 依赖于其与巨噬细胞表面的 PLXDC2 相互作用来激活抗肿瘤免疫并抑制肿瘤生长。
06、PTEN-PLXDC2 参与激活 JAK2-STAT1 信号以重新编程巨噬细胞
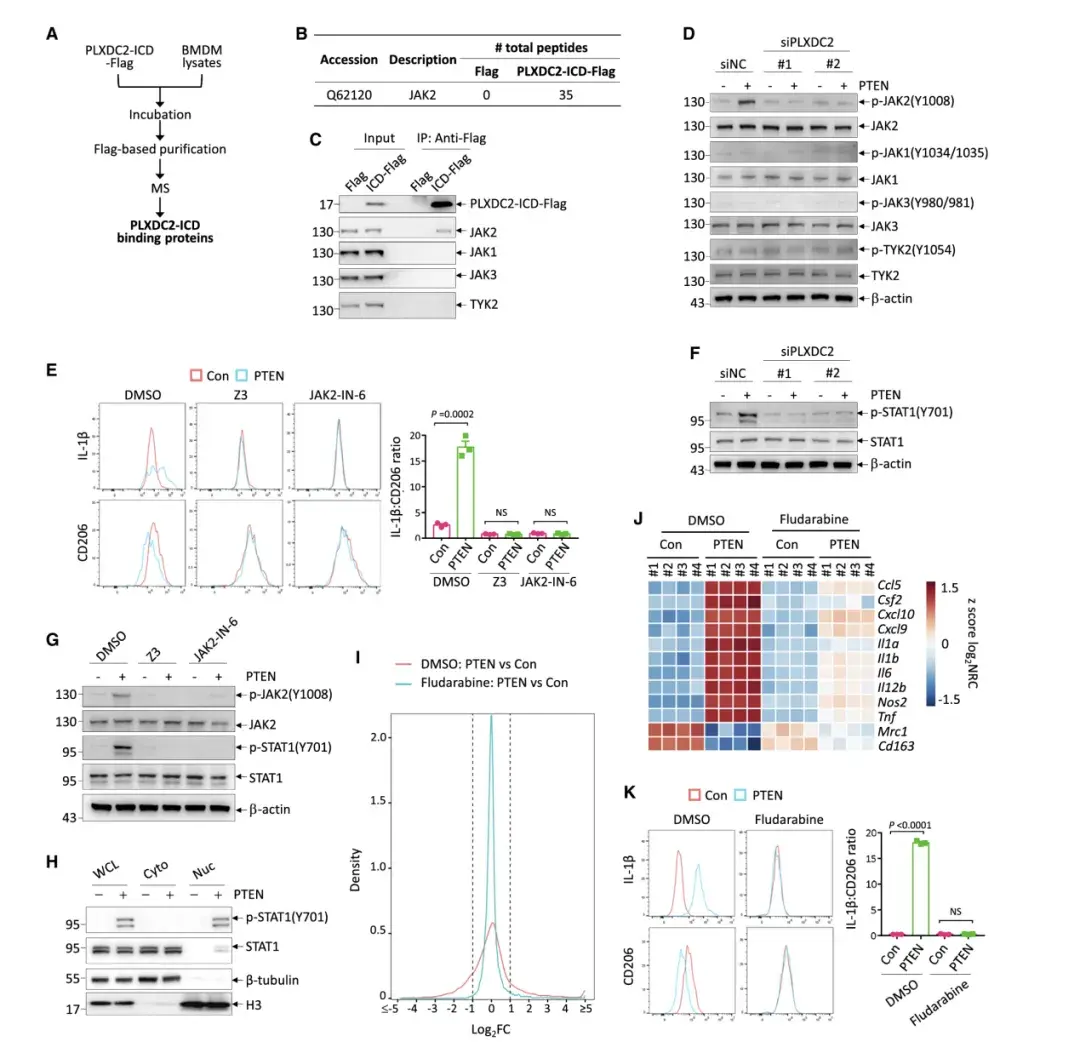
PLXDC2的下游信号传导在很大程度上是未知的。作者发现在所有已确定与PLXDC2-ICD相互作用的蛋白质中,Janus 激酶 2 (JAK2) 表现出最强的强度。作者验证了PLXDC2-ICD与 BMDM中的 JAK2而非其他家族成员的选择性结合以及全长PLXDC2与293T细胞中异位表达的JAK2之间的相互作用。
试验验证了PLXDC2-ICD与JAK2的选择性结合,并且PTEN处理增加了M2极化巨噬细胞中JAK2的磷酸化,这一过程依赖于PLXDC2。使用JAK2特异性抑制剂发现,它们能阻断PTEN在巨噬细胞极化中的作用,表明PLXDC2通过PTEN激活JAK2来调节巨噬细胞极化。
TEN处理以PLXDC2和JAK2依赖的方式特异性地增加了 STAT1 (Tyr701) 的磷酸化,但未增加其他 STAT的磷酸化,表明PTEN PLXDC2-JAK2轴激活了STAT1。STAT1在巨噬细胞的M1极化中起着既定的作用。事实上,STAT1 抑制表现出与PLXDC2 敲低类似的效果,可拮抗细胞外PTEN对巨噬细胞重编程的影响。总的来说,作者确定了JAK2-STAT1信号传导负责介导巨噬细胞中PTEN-PLXDC2的下游信号传导。
07、细胞外PTEN与PD-1阻断协同作用以抑制肿瘤生长
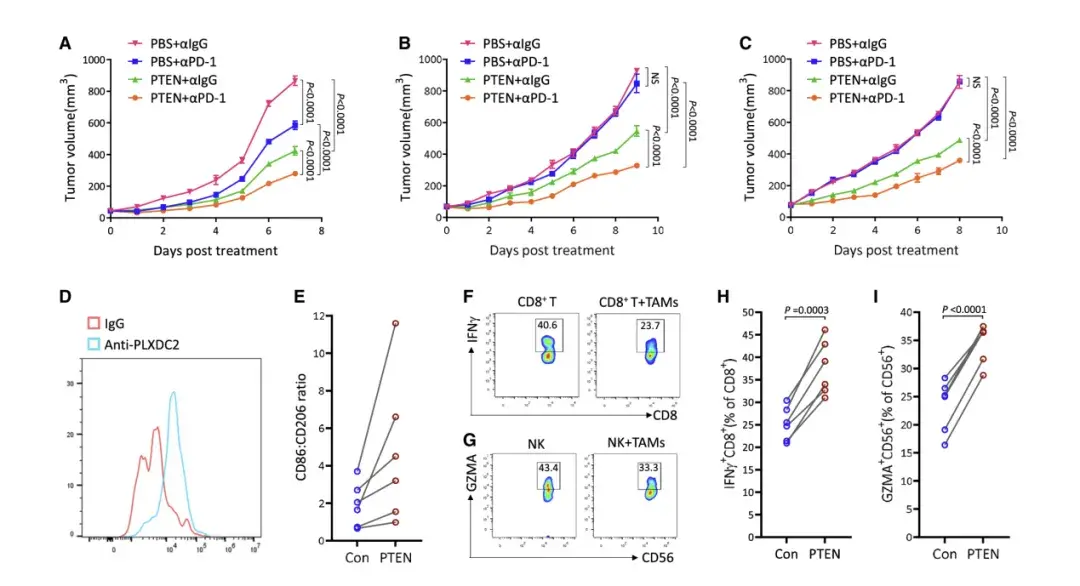
研究表明,M2 样TAM对各种治疗剂产生耐药性,如PD-1阻断。因此,作者测试了 PTEN 诱导的促肿瘤发生M2样TAM 重编程为抗肿瘤 M1 样表型是否会导致对免疫检查点阻断的反应增强。单用PTEN 或 PD-1 阻断抗体治疗都可缩小 B16-F10-DPTEN 肿瘤的大小,而联合 PTEN/抗 PD-1 治疗显示出最显著的效果。
PD-1阻断不影响细胞外PTEN水平或TAM上PLXDC2表达,表明其作用独立于PTEN-PLXDC2轴。在CT26结直肠癌和Lewis肺癌模型中,PD-1阻断对肿瘤负荷无影响,但PTEN治疗显示出中等抗肿瘤作用,联合治疗则显著减小了肿瘤大小。总体而言,PTEN治疗增强了体内抗PD-1治疗的效果。
为了解 PLXDC2在 TAM 中的临床意义,作者创建了两个基因特征来代表 PLXDC2 + 和 PLXDC2 TAM,并将它们应用于TCGA 数据库中的大量 RNA-seq 数据集,使用 CIBERSORTx 工具来确定每个样本中 PLXDC2 + 和 PLXDC2 TAM 的比例。分析显示,PLXDC2 + TAM 比例较高的患者总生存率较低。在TCGA 数据库的COAD、ESCA 和 LIHC 患者中也观察到类似结果。这些发现与骨髓特异性 Plxdc2耗尽小鼠肿瘤生长延迟的结果一致。
从结肠直肠癌患者的肿瘤组织中分离出肿瘤浸润性TAM,并确认其表达 PLXDC2。作者发现,所有患者的 TAM 在接受 PTEN 治疗后,CD86 与 CD206 的表达比率均增加,表明 PTEN 诱导患者来源的 TAM 的 M2 极化至 M1 极化。作者还从每位患者的外周血中分离出 CD8+ T 和NK 细胞,并使用来自一名患者的样本示例性地证实了自体 TAM 抑制了这些细胞。然后,每个患者的TAM经PTEN处理后与自体CD8+ T和NK细胞孵育。结果表明,与 PTEN 处理的 TAM 一起孵育的 CD8+ T 和 NK 细胞比与未用 PTEN 处理的 TAM 一起培养的 CD8+ T 和 NK 细胞表现出更高的活性,表明 PTEN 在减轻患者来源的 TAM 的免疫抑制作用方面发挥作用。因此,体外数据表明,患者活组织检查的TAM对PTEN治疗有反应。
派森诺目前已致力于单细胞测序多年,凭借其丰富的项目经验、出色的服务能力和在疑难样本解离方面独特经验,赢得了业界的广泛赞誉。目前,派森诺生物单细胞转录组测序极速周期活动正在火热进行中,详情请联系当地销售。